






Potential Relationship between Extremophiles and Hydrocarbon Resources in Marine Extreme Environment
-
摘要: 为了弄清海洋极端环境下微生物参与油气资源形成和演化的潜在机制, 进行了现代海洋热泉和冷泉等环境中微生物类型分析和生物量估算, 探讨了极端微生物活动和油气资源的潜在关系.认为海洋极端环境下微生物类型主要为细菌和古细菌, 热泉微生物群落主要为异养发酵菌、硫酸盐还原菌、产甲烷菌等; 冷泉微生物群落主要为ANME-2族的厌氧甲烷氧化古细菌、硫酸盐还原细菌和ANME-1族厌氧甲烷氧化古菌.这些极端微生物利用CH4和H2S等气体进行能量固定, 有较高的生物丰度和较低的分异度, 具有垂向和水平分带性, 并能营生一套独特的宏体生物.极端微生物活动直接和间接地参与了油气资源的形成和改造, 示踪海底油气资源的变迁.对于探索地球早期海洋微生物活动与油气资源形成, 寻找地史时期或华南地史早期烃源岩具有重要理论和实践指导意义.Abstract: To understand the potential mechanism of marine extremophiles participating in the formation and the evolution of hydrocarbon resources in marine extreme environments, some typical kinds of extremophiles and their distributions in marine hydrothermal venting and cold venting are discussed and evaluated respectively. The potential relationship between extremophile activities and hydrocarbon resources in marine extreme environments are then discussed in details. It could be now preliminary concluded that archaea and bacteria are the two main kinds of extremophiles in marine extreme environments. The dominating microbial communities in hydrothermal venting are heterotrophic zymogens, sulphate reducers and methanogens, while the ANME-2 group (Methanosarcinales) surrounded by sulfate-reducing bacteria and ANME-1 group are dominated in cold venting. Marine extremophiles would be able to use CH4 and H2S to synthesize energy for metabolism and to support food chains for other unique macrobiota nearby, which together present a high abundance but a low diversity with distinct characteristics of horizontal and vertical distributions. Marine extremophiles might play an important role either directly or indirectly in the processes of hydrocarbon formation and later alteration, and could indicate the evolution of hydrocarbon resources in marine extreme environments. Our research thus has a great significance both in theoretical approach of potential hydrocarbon resources formed by marine extremophile activities and in practical exploration of the potential hydrocarbon source sedimentary layers formed in the earth history or the potential strata in South China.
-
Baross, J. A., Hoff man, S. E., 1985. Submarine hydrothermalvents and associated gradient environments as sites for the origin and evolution of life. Origins Life, 15: 327-345. Boetius, A., Suess, E., 2004. Hydrate ridge: A natural labo-ratory for the study of microbial life fueled by methane from near-surface gas hydrates. Chemical Geology, 205: 291-310. doi: 10.1016/j.chemgeo.2003.12.034 Bons, P. D., Montenari, M., Hartmann, D., et al., 2004. Fos-sil microbes in late Proterozoic fibrous calcite veins from Arkaroola, South Australia. In: Geological Society of America, 2004 annual meeting, Anonymous. Ab-stracts with Programs-Geological Society of America (November 2004), 36 (5): 475. Brasier, M. D., Green, O. R., Jephcoat, A. P., et al., 2002. Questioning the evidence for earth′s oldest fossils. Na-ture, 416: 76-81. Chen, J., Lian, B., Wang, B., et al., 2006. The occurrence and biogeochemistry of microbes in extreme environ-ments. Earth Science Frontiers, 13 (6): 199-207 (in Chinese with English abstract). Daffonchio1, D., Borin1, S., Brusa1, T., et al., 2006. Strati-fied prokaryote network in the oxic-anoxic transition of a deep-sea halocline. Nature, 440: 203-207. doi: 10.1038/nature04418 Dang, H. Y., Li, T. G., Zeng, Z. G., et al., 2006. Microbio-logical studies on sub-sea floor deep biosphere in deep sea extreme environments. Studia Marina Sinica, 47: 41-60 (in Chinese with English abstract). Deming, J. W., Baross, J. A., 1993. Deep-sea smokers: Win-dows to a sub-surface biosphere. Geochimicaet Cosmo-chimica Acta, 57: 3219-3229. doi: 10.1016/0016-7037(93)90535-5 Dutkiewicz, A., Volk, H., George, S., et al., 2006. Biomark-ers from Huronian oil-bearing fluid inclusions: An un-contaminated record of life before the Great Oxidation Event. Geology, 34 (6): 437-440. doi: 10.1130/G22360.1 Dutkiewicz, A., Volk, H., Ridley, J., et al., 2003. Biomark-ers, brines, and oil in the Mesoproterozoic, Roper Super-basin, Australia. Geology, 31 (11): 981-984. doi: 10.1130/G19754.1 Feng, J., Li, J. H., Niu, X. L., 2005. Research advances in hydrothermal vent microbial communities and its signif-icance for geology. Advances in Earth Science, 20 (7): 732-739 (in Chinese with English abstract). Fink, C. R., Spence, G. D., 1999. Hydrate distribution off Vancouver Island from multifrequency single-channel seismic reflection data. J. Geophys. Res. (Solid Earth), 104: 2909-2922. doi: 10.1029/98JB02641 Furnes, H., Banerjee, N. R., Muehlenbachs, K., et al., 2004. Early life recorded in archean pillow lavas. Science, 304 (5670): 578-581. doi: 10.1126/science.1095858 Hinrichs, K. U., Hayes, J. M., Sylva, S. P., et al., 1999. Methane-consuming archaebacteria in marine sedi-ments. Nature, 398 (6730): 802-805. doi: 10.1038/19751 Horita, J., Berndt, M. E., 1999. Abiogenic methane forma-tion and isotopic fractionation under hydrothermal con-ditions. Science, 285: 1055-1057. doi: 10.1126/science.285.5430.1055 Ian, M., Head, D., Martin, J., et al., 2003. Biological activityin the deep subsurface and the origin of heavy oil. Na-ture, 344-352. IPSC, 2001. Earth, oceans and life: Integrated ocean drilling program, initial science plan, 2003-2013. IWG Sup-porting Office, Washington DC, 110. Jiang, G. Q., Kennedy, M. J., Christie-Blick, N., 2003. Stableisotopic evidence for methane seeps in Neoproterozoic postglacial cap carbonates. Nature, 426: 822-826. doi: 10.1038/nature02201 Jiang, G. Q., Kennedy, M. J., Christie-Blick, N., et al., 2006. Stratigraphy, sedimentary structures, and textures of the late Neoproterozoic Doushantuo cap carbonate in South China. Journal of Sedimentary Research, 76: 978-995. doi: 10.2110/jsr.2006.086 Krajick, K., 2007. Race to plumb the frigid depths. Science, 315: 1525-1528. doi: 10.1126/science.315.5818.1525 Krejci-Graf, K., 1932. Rule of density of oils. Bull. Am. As-soc. Petrol. Geol. , 16: 1038. Kruglyakova, R. P., Byakov, Y. A., Kruglyakova, M. V., et al., 2004. Natural oil and gas seeps on the Black Sea floor. Geo-Mar. Lett. , 24: 150-162. Li, Q., Wang, J. S., Chen, Q., et al., 2006. Barite within the cap dolomite in Three Gorges area and its palaeogeo-graphic meaning. Journal of Northwest University (Natural Science Edition), 36 (special issue): 196-200 (in Chinese with English abstract). Li, Y. L., Peacock, A. D., White D. C., et al., 2007. Spatial patterns of bacterial signature biomarkers in marine sediments of the Gulf of Mexico. Chemical Geology, 238: 168-179. doi: 10.1016/j.chemgeo.2006.11.007 Liu, J. F., Mu, B. Z., 2004. Extreme environment of oil reser-voir and associated microorganisms. Journal of Micro-biology, 24 (4): 31-34 (in Chinese with English ab-stract). Lollar, S. B., Westgate, T. D., Ward, J. A., et al., 2002. Abiogenic formation of alkanes in the earth′s crust as a minor source for global hydrocarbon reservoirs. Na-ture, 416: 522-524. Margesin, R., Schinner, F., 2001. Biodegradation and biore-mediation of hydrocarbons in extreme environments. Appl. Microbiol. Biotechnol. , 56: 650-663. doi: 10.1007/s002530100701 Marsh, A. G., Mullineaux, L. S., Young, C. M., et al., 2001. Larval dispersal potential of the tubeworm Riftia pachy-ptila at deep-sea hydrothermal vents. Nature, 411: 77-80. doi: 10.1038/35075063 Nussbaumer, A. D., Fisher, C. R., Bright, M., 2006. Hori-zontal endosymbiont transmission in hydrothermal venttubeworms. Nature, 441: 345-348. doi: 10.1038/nature04793 Orphan, V. J., House, C. H., Hinrichs, K. U., et al., 2001. Methane-consuming archaea revealed by directly cou-pled isotopic and phylogenetic analysis. Science, 293 (5529): 484-487. doi: 10.1126/science.1061338 Orphan, V. J., House, C. H., Hinrichs, K. U., et al., 2002. Multiple archaeal groups mediate methane oxidation in anoxic cold seep sediments. Proceedings of the National Academy of Sciences of the United States of America, 99 (11): 7663-7668. doi: 10.1073/pnas.072210299 Parkes, R. J., Cragg, B. A., Wellsbury, P., 2000. Recentstudies on bacterial populations and processes in subseafloor sediments: A review. Hydrogeological Jour-nal, 8 (1): 11-28. doi: 10.1007/PL00010971 Paull, C. K., Hecker, B., Commeau, R., et al., 1984. Biologi-cal communities at the Florida escarpment resemble hy-drothermal vent taxa. Science, 226: 965-967. doi: 10.1126/science.226.4677.965 Paull, C. K., Schlining, B., Ussler Ⅲ, W., et al., 2005. Dis-tribution of chemosynthetic biological communities in Monterey Bay, California. Geology, 33 (2): 85-88. doi: 10.1130/G20927.1 Pradillon, F., Shillito, B., Young, C. M., et al., 2001. Devel-opmental arrest in vent worm embryos. Nature, 413: 698-699. doi: 10.1038/35099674 Prieur, D., 1997. Microbiology of deep-sea hydrothermalvents. Marine Biotechnology, 15: 42-244. Rasmussen, B., 2005. Evidence for pervasive petroleum gen-eration and migration in 3.2 and 2.63 Ga shales. Geolo-gy, 497-500. Rasmussen, B., Buick, R., 2000. Oil old ores: Evidence for hydrothermal petroleum generation in an Archean vol-canogenic massive sulfide deposit. Geology, 731-734. Rothschild, L. J., Mancinelli, R. L., 2001. Life in extreme en-vironments. Nature, 409: 1092-1101. doi: 10.1038/35059215 Rusanov, I. I., Lein, A. Y., Pi menov, N. V., et al., 2002. The biogeochemical cycles of methane on the northwesternshelf of the Black Sea. Microbiology, 71 (4): 479-487. doi: 10.1023/A:1019862014508 Science Committee of ODP China, 2003. Chinese national sci-ence plan (2003-2013) for participation in IODP. Ad-vances in Earth Science, 18 (5): 662-665 (in Chinese with English abstract). Seewald, J. S., 2003. Organic-inorganic interactions in petro-leum-producing sedimentary basins. Nature, 426: 327-333. doi: 10.1038/nature02132 Si moneit, B. R. T., Lonsdale, P. F., 1982. Hydrothermal pe-troleum in mineralized mounds at the seabed of Guay-mas basin. Nature, 295: 198-202. doi: 10.1038/295198a0 Su, X., Chen, F., Wei, S. P., et al., 2007. Preliminary study on the correlation between microbial abundance and methane concentration in sediments from cold seeps in the northern South China Sea. Geoscience, 21 (1): 101-104 (in Chinese with English abstract). Takai, K., Gamo, T., Tsunogai, U., et al., 2004. Geochemi-cal and microbiological evidence for a hydrogen-based, hyperthermophilic subsurface lithoautotrophic microbial ecosystem (HyperSLME) beneath an active deep-sea hydrothermal field. Extremophiles, 8 (4): 269-282. doi: 10.1007/s00792-004-0386-3 Tarasov, V. G., Gebruk, A. V., Mironov, A. N., et al., 2005. Deep-sea and shallow-water hydrothermal ventcommunities: Two different phenomena? Chemical Ge-ology, 224: 5-39. Wang, J. S., Gan, H. Y., Wei, Q., et al., 2005. Stable iso-topes of carbon and sulfur of cap dolomite in the Three Gorges and its mechanism discussion. Geoscience, 19 (1): 14-20 (in Chinese with English abstract). Wang, P. X., 2003. Earth system science in China Quo Va-dis? Advances in Earth Science, 18 (6): 837-851 (in Chinese with English abstract). Wang, W. C., Tao, M. X., 2005. Geomicrobial processes and petroleum resources. Geological Bulletin of China, 24 (10-11): 1022-1026 (in Chinese with English ab-stract). Wilhel ms, A., Larter, S. R., Head, I., et al., 2001. Biodegra-dation of oil in uplifted basins prevented by deep-burial sterilization. Nature, 411: 1034-1037. doi: 10.1038/35082535 Wilson, H. H., 1990. The case for early generation and accu-mulation of oil. Journal of Petroleum Geology, 13 (2): 127-156. doi: 10.1111/j.1747-5457.1990.tb00836.x Yang, R. D., Zhu, L. J., Gao, H., et al., 2005. A study on characteristics of the hydrothermal vent and relating bi-ota at the Cambrian bottom in Songlin, Zunyi County, Guizhou Province. Geological Review, 51 (5): 481-492 (in Chinese with English abstract). You, C. F., Bickle, M. J., 1998. Evolution of an active sea-floor massive sulphide deposit. Nature, 394: 668-671. doi: 10.1038/29279 Zhang, C. L., Lanoil, B., 2004. Geomicrobiology and biogeo-chemistry of gas hydrates and cold seeps. Chemical Ge-ology, 205 (3-4): 187-194. Zhang, M., Dong, X. Z., 2006. A survey of extromophiles project supported by 973. Acta Microbiologica Sinica, 46 (2): 336 (in Chinese). Zhou, Q., Du, Y. S., Wang, J. S., et al., 2007. The character-istics and significance of the cold seep carbonates from Datangpo Formation, Nanhua Serise in the northeast of Guizhou. Earth Science—Journal of China University of Geoscicenes, 32 (3): 339-346 (in Chinese with Eng-lish abstract). 陈骏, 连宾, 王斌, 等, 2006. 极端环境下的微生物及其生物地球化学作用. 地学前缘, 13 (6): 199-207. doi: 10.3321/j.issn:1005-2321.2006.06.023 党宏月, 李铁刚, 曾志刚, 等, 2006. 深海极端环境深部生物圈微生物学研究综述. 海洋科学集刊, 47: 41-60. https://www.cnki.com.cn/Article/CJFDTOTAL-HKJK200600003.htm 冯军, 李江海, 牛向龙, 2005. 现代海底热液微生物群落及其地质意义. 地球科学进展, 20 (7): 732-739. doi: 10.3321/j.issn:1001-8166.2005.07.005 李清, 王家生, 陈祈, 等, 2006. 三峡"盖帽"白云岩中重晶石研究及其古地理意义. 西北大学学报(自然科学版), 36 (专辑): 196-200. 刘金峰, 牟伯中, 2004. 油藏极端环境中的微生物. 微生物学杂志, 24 (4): 31-34. doi: 10.3969/j.issn.1005-7021.2004.04.011 苏新, 陈芳, 魏士平, 等, 2007. 南海北部冷泉区沉积物中微生物丰度与甲烷浓度变化关系的初步研究. 现代地质, 21 (1): 101-104. doi: 10.3969/j.issn.1000-8527.2007.01.012 王家生, 甘华阳, 魏清, 等, 2005. 三峡"盖帽"白云岩的碳、硫稳定同位素研究及其成因探讨. 现代地质, 19 (1): 14-20. doi: 10.3969/j.issn.1000-8527.2005.01.002 汪品先, 2003. 我国的地球系统科学研究向何处去. 地球科学进展, 18 (6): 837-851. doi: 10.3321/j.issn:1001-8166.2003.06.003 王万春, 陶明信, 2005. 地质微生物作用与油气资源. 地质通报, 24 (10-11): 1022-1026. https://www.cnki.com.cn/Article/CJFDTOTAL-ZQYD2005Z1026.htm 杨瑞东, 朱立军, 高慧, 等, 2005. 贵州遵义松林寒武系底部热液喷口及与喷口相关生物群特征. 地质论评, 51 (5): 481-492. doi: 10.3321/j.issn:0371-5736.2005.05.001 张敏, 东秀珠, 2006.973项目"极端微生物及其功能利用的基础研究"研究进展. 微生物学报, 46 (2): 336. doi: 10.3321/j.issn:0001-6209.2006.02.035 中国大洋钻探学术委员会, 2003. 中国加入综合大洋钻探(IODP) 科学计划(2003-2013). 地球科学进展, 18 (5): 662-665. doi: 10.3321/j.issn:1001-8166.2003.05.004 周琦, 杜远生, 王家生等, 2007. 黔东北地区南华系大塘坡组冷泉碳酸盐岩及其意义. 地球科学——中国地质大学学报, 32 (3): 339-346. https://www.cnki.com.cn/Article/CJFDTOTAL-DQKX200703005.htm -
 下载:
下载:
 点击查看大图
点击查看大图
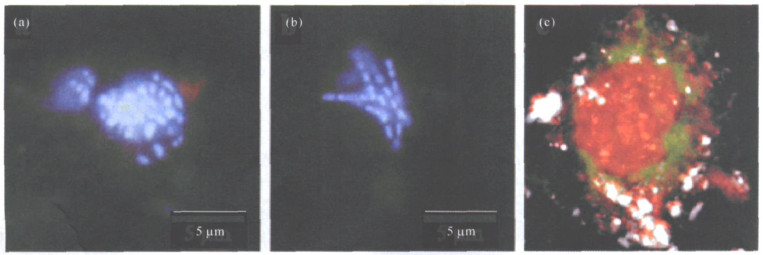
图(1)
计量
- 文章访问数: 3472
- HTML全文浏览量: 833
- PDF下载量: 224
- 被引次数: 0
