






Calci-Microbialite as a Kind of Potential Hydrocarbon Source Rock and Its Geomicrobiological Processes
-
摘要: 钙质微生物岩是各种微生物生命活动引起的特殊的碳酸盐建造, 主要集中发育在前寒武纪及显生宙等重大地质转折期, 在我国华北及华南地区有着广泛的分布.微生物岩中大量微生物化石的存在反映当时的海洋具有很高的初级生产力.明显偏低的钼同位素值显示在前寒武纪相当长的时期里, 海洋底部处于缺氧状态.Th/U值的急剧降低和黄铁矿在浅水台地区的广泛出现显示二叠纪大绝灭后, 海洋环境也同样呈现缺氧的还原状态.高生产力和还原的沉积环境为生物有机质的埋藏及烃源岩的形成创造了条件.前寒武纪及晚泥盆世微生物岩中沥青的发现, 进一步增加了钙质微生物岩作为潜在碳酸盐型烃源岩的可能性.Abstract: Calci-microbialite is a special carbonate buildup, formed due to the activities of different kinds of microbes. Most of microbialites were developed in Precambrian or at the great transitional stages in Phanerozoic. Widespread microbialites have been found both in North and South China. Abundant microfossils preserved in the microbialites show high-level productivity during deposition. The obviously lower value of Mo isotope shows that the seafloor is of anoxic condition during the long time of Precambrian. The sharp reduction of Th/U value as well as the wide spread pyrite in shallow carbonate platforms indicate that the ocean environment is anoxic after end-Permian mass extinction. High-level productivity and anoxic sedimentary environment favor the efficient preservation of organic matter and thus the formation of source rocks. Bitumen observed in many outcrops of Precambrian and Late Devonian microbialites further raises the possibility of calci-microbialite as a potential hydrocarbon source rock.
-
Key words:
- calci-microbialite /
- potential hydrocarbon source rock /
- geomicrobes
-
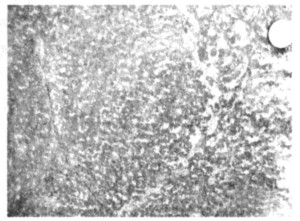
图 1 重庆老龙洞二叠纪-三叠纪之交钙质微生物岩的“花斑状”构造, 照片中白色部分由微晶碳酸盐矿物所组成, 暗色部分由中粗晶碳酸盐矿物所组成
Fig. 1. "Graniphyic fabric" structure of the P/T boundary microbialites in Laolongdong section, Chongqing. The white part of the picture was composed of microlite carbonate minerals, the darkness composed of mid-coarse carbonate minerals
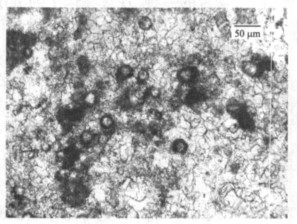
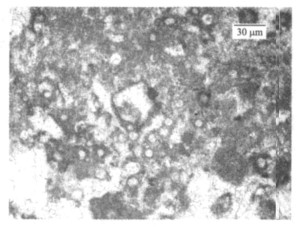
图 3 二叠纪-三叠纪之交钙质微生物岩中的微生物化石
Fig. 3. Microfossils in the calci-microbialite near the Permian-Triassic boundary
-
Arnold, G. L., Anbar, A. D., Barling, J., et al., 2004. Molyb-denum isotope evidence for widespread anoxia in mid-Proterozoic oceans. Science, 304 (5667): 87-90. doi: 10.1126/science.1091785 Baud, A., Cirilli, S., Marcoux, J., 1997. Biotic response to mass extinction: The lowermost Triassic microbialites. Facies, 36: 238-242. Baud, A., Richoz, S., Marcoux, J., 2005. Calci microbial caprocks from the basal Triassic units of the Taurus (SW Turkey), an anachronistic facies before the biotic recov-ery. Comptes Rendus. Palevol. , 4 (6-7): 569-582. doi: 10.1016/j.crpv.2005.03.001 Baud, A., Richoz, S., Pruss, S. B., 2007. The lower Triassic anachronistic carbonate facies in space and time. Globaland Planetary Change, 55 (1-3): 81-89. doi: 10.1016/j.gloplacha.2006.06.008 Burne, R. V., Moore, I. S., 1987. Microbialites: Organosedi-mentary deposits of benthic microbial communities. Palaios, 2 (3): 241-254. doi: 10.2307/3514674 Cao, R. J., Yuan, X. L., 2003. Brief history and current status of stromatolite study in China. Acta Micropalaeontologica Sinica, 20 (1): 5-14 (in Chinese with English abstract). Chen, J. B., 1993. Progress and problems in research on stro-matolites. In: Zhu, S. X., et al., eds., The stromatolites of China. Tianjin Univ. Press, Tianjin, 205-214 (in Chinese with English abstract). Dai, Y. D., Chen, M. E., Wang, Y., 1996. Development and per-spective of research for microbialites. Advances in Earth Sci-ence, 11 (2): 209-215 (in Chinese with English abstract). Dupraz, C., Visscher, P. T., Baumgartner, L. K., et al., 2004. Microbe-mineral interactions: Early carbonate precipitation in a hypersaline lake (Eleuthera Island, Ba-hamas). Sedimentology, 51: 745-765. doi: 10.1111/j.1365-3091.2004.00649.x Heydari, E., Hassanzadeh, J., Wade, W. J., et al., 2003. Per-mian-Triassic boundary interval in the Abadeh section of Iran with implications for mass extinction: Part 1. Sedimentology. Palaeogeography, Palaeoclimatology, Palaeoecology, 193 (3-4): 405-423. doi: 10.1016/S0031-0182(03)00258-X Hips, K., Haas, J., 2006. Calcimicrobial stromatolites at the Permian-Triassic boundary in a western Tethyan sec-tion, Bükk Mountains, Hungary. Sedimentary Geology, 185: 239-253. doi: 10.1016/j.sedgeo.2005.12.016 Kershaw, S., Zhang, J., Lan, G., 1999. A microbialite carbonate crust at the Permian-Triassic boundary in South China, and its palaeoenvironmental significance. Palaeogeography, Palaeoclimatology, Palaeoecology, 146: 1-18. doi: 10.1016/S0031-0182(98)00139-4 Konishi, Y., Prince, J., Knott, B., 2001. The fauna of throm-bolitic microbialites, Lake Clifton, Western Australia. Hydrobiologia, 457 (1-3): 39-47. Lehrmann, D. J., 1999. Early Triassic calcimicrobial mounds and biostromes of the Nanpanjiang basin, South China. Geology, 27 (4): 359-362. doi: 10.1130/0091-7613(1999)027<0359:ETCMAB>2.3.CO;2 Li, Y. C., Zhou, Z. Z., 2002. Massive dissociation of gas hydrateduring oceanic anoxia as a cause of mass extinction at the end of Permian. Geology Geochemistry, 30 (1): 57-63 (inChinese with English abstract). Liang, Y. Z., Cao, R. J., Zhang, L. Y., et al., 1984. Pseud-ogymnosolenaceae of Late Precambrian in China. Geo-logical Publishing House, Beijing, 1-200 (in Chinese). Liang, Y. Z., Wang, Y., Du, R. L., 1993. Structural deformation of microstromatolite in the Bayan Obo Group of Inner Mongolia and its geological significance. Geological Bulletin of China, 3: 229-238 (in Chinese with English abstract). Liang, Y. Z., Zhu, S. X., Gao, Z. J., et al., 1995. New progress in the study of stratomalites—microbialite. Regional Geology of China, 1: 57-65 (in Chinese with English abstract). Marcoux, J., Baud, A., 1986. The Permo-Triassic boundary in the Antalya nappes (western Taurides, Turkey). Me-moria della Societa Geologica Italiana, 34: 243-252. Qiu, S. Y., Liang, Y. Z., Cao, R. J., et al., 1992. Late Pre-cambrian stromatolites and related mineral product. Northwest University Press, Xi′an (in Chinese). Riding, R., Liang, L. Y., 2005. Geobiology of microbial carbon-ates: Metazoan and seawater saturation state influences onsecular trends during the Phanerozoic. Palaeogeography, Palaeoclimatology, Palaeoecology, 219: 101-115. doi: 10.1016/j.palaeo.2004.11.018 Sano, H., Nakashima, K., 1997. Lowermost Triassic (Gries-bachian) microbial bindstone-cementstone, Southwest Japan. Facies, 36: 1-24. doi: 10.1007/BF02536874 Sheehan, P. M., Harris, M. T., 2004. Microbialite resurgence af-ter the Late Ordovician extinction. Nature, 430: 75-78. doi: 10.1038/nature02654 Shen, J. W., Webb, G. E., 2004a. Famennian (Upper Devoni-an) calcimicrobial (Renalcis) reef at Miaomen, Guilin, Guangxi, South China. Palaeogeography, Palaeoclima-tology, Palaeoecology, 204: 373-394. doi: 10.1016/S0031-0182(03)00737-5 Shen, J. W., Webb, G. E., 2004b. Famennian (Upper Devonian) stromatolite reefs at Shatang, Guilin, Guangxi, South Chi-na. Sedimentary Geology, 170 (1-2): 63-84. doi: 10.1016/j.sedgeo.2004.06.004 [84] Walter, M. R., 1994. Stromatolites: The main geological source of information on the evolution of early benthos. In: Bengtson, S., ed., Early life on earth. Columbia U. P., New York, 270-286. Wang, Y. B., Tong, J. N., Wang, J. S., et al., 2005. Calcimicrobialite after end-Permian mass extinctionin South China and its palaeoenvir-onmental significance. Chinese Science Bulletin, 50: 665-671. doi: 10.1360/982004-323 Wignall, P. B., Twitchett, R. J., 1996. Oceanic anoxia and the end Permian mass extinction. Science, 272: 1155-1158. doi: 10.1126/science.272.5265.1155 Wignall, P. B., Twitchett, R. J., 1999. Unusual intraclastic limestones in Lower Triassic carbonates and their bear-ing on the aftermath of the end-Permian mass extinc-tion. Sedimentology, 46 (2): 303-316. doi: 10.1046/j.1365-3091.1999.00214.x Wignall, P. B., Twitchett, R. J., 2002. Permian-Triassic sedi-mentology of Jameson Land, East Greenland: Incised submarine channels in an anoxic basin. Journal of the Geological Society, 159: 691-703. doi: 10.1144/0016-764900-120 Xie, S. C., Gong, Y. M., Tong, J. N., et al., 2006. Advance-ment from paleontology to geobiology. Chinese Science Bulletin, 51 (19): 2327-2336 (in Chinese). doi: 10.1007/s11434-006-2111-3 Xie, S. C., Pancost R. D., Yin H. F., et al., 2005. Two epi-sodes of microbial change coupled with Permo/Triassic faunal mass extinction. Nature, 434: 494-497. doi: 10.1038/nature03396 Yan, X. Q., Meng, F. W., Yuan, X. L., 2006. Geochemical characteristics of the cherts of the Neoproterozoic Jiud-ingshan Formation in northern Jiangsu and Anhui prov-inces. Acta Micropalaeontologica Sinica, 23 (3): 295-302 (in Chinese with English abstract). Zhu, S. X., et al., 1993. The stromatolites of China. Tianjin Univ. Press, Tianjin, 1-263 (in Chinese). 曹瑞骥, 袁训来, 2003. 中国叠层石研究的历史和现状. 微体古生物学报, 20 (1): 5-14. doi: 10.3969/j.issn.1000-0674.2003.01.002 [104] 陈晋镳, 1993. 叠层石研究的进展和问题. 见: 朱士兴, 等, 中国叠层石. 天津: 天津大学出版社, 205-214. 戴永定, 陈孟莪, 王尧, 1996. 微生物岩研究的发展与展望. 地球科学进展, 11 (2): 209-215. doi: 10.3321/j.issn:1001-8166.1996.02.016 李玉成, 周忠泽, 2002. 华南二叠纪末缺氧海水中的有毒气体与生物集群绝灭. 地质地球化学, 30 (1): 57-63. doi: 10.3969/j.issn.1672-9250.2002.01.010 梁玉左, 曹瑞骥, 张录易, 等, 1984. 晚前寒武纪假裸枝叠层石. 北京: 地质出版社. 梁玉左, 王楫, 杜汝霖, 1993. 内蒙古白云鄂博群中微小型叠层石(Ministrmatilites) 的构造变形及其地质意义. 地质通报, 3: 229-238. https://www.cnki.com.cn/Article/CJFDTOTAL-ZQYD199303005.htm 梁玉左, 朱士兴, 高振家, 等, 1995. 叠层石研究的新进展——微生物岩. 中国区域地质, 1: 57-65. https://www.cnki.com.cn/Article/CJFDTOTAL-ZQYD501.008.htm 邱树玉, 梁玉左, 曹瑞骥, 等, 1992. 晚前寒武纪叠层石及相关矿产. 西安: 西北大学出版社. 谢树成, 龚一鸣, 童金南, 等, 2006. 从古生物学到地球生物学的跨越. 科学通报, 51 (19): 2327-2336. doi: 10.3321/j.issn:0023-074X.2006.19.018 严贤勤, 孟凡巍, 袁训来, 2006. 徐淮地区新元古代九顶山组燧石结核的地球化学特征. 微体古生物学报, 23 (3): 295-302. doi: 10.3969/j.issn.1000-0674.2006.03.005 朱士兴, 等, 1993. 中国叠层石. 天津: 天津大学出版社, 1-263. -
 下载:
下载:

 点击查看大图
点击查看大图
图(4)
计量
- 文章访问数: 3759
- HTML全文浏览量: 681
- PDF下载量: 167
- 被引次数: 0
