






Vegetation and Climate Changes around Celebes Sea during Holocene
-
摘要: 全新世时期的环境和气候变化是全球气候模拟、预测中不可或缺的资料.对苏拉威西海西北部MD98-2178孔(3.6200°N,118.7000°E,水深1 984 m)全新世的样品进行孢粉分析和浮游有孔虫氧稳定同位素测试,重建全新世苏拉威西海周边地区植被演化和气候变化图景.根据孢粉记录得到:在全新世早期至7 ka BP时,各孢粉组合浓度剧烈下降,指示海平面处于上升阶段;在7~4 ka BP时,各孢粉组合浓度都处于低谷,体现为高海平面期;在4 ka BP之后,孢粉记录则显示海平面有小幅的下降.在全新世中期,即5~4 ka BP,热带高山雨林花粉含量明显上升,表明是温度低值期.蕨类孢子记录显示降雨量在全新世早期是持续增加的,但在全新世中期之后,降雨量有所减少,这与陆地孢粉记录和印尼石笋记录的结果相似,体现全新世该地区降雨量受海陆格局和太阳活动共同影响.Abstract: The environment and climate information during the Holocene is indispensable data for the global climate modeling and forecasting. Scenarios of regional vegetation and climate changes around the Celebes Sea during the Holocene are explored based on the alynological and oxygen isotope records of core MD98-2178 (3.6200°N, 118.7000°E; 1 984 m water depth) from the northwest Celebes Sea in this study. Sea-level changes are discussed since the marine pollen record of core MD98-2178 is considered as an indicator. The sharp decline in concentration of all pollen groups during the Early Holocene, ~10-7 ka BP, indicates a sea-level rise. All pollen groups at 7-4 ka BP suggest sustaining of a high sea-level. The marine pollen record after 4 ka BP shows a slight fall of sea-level. The significant increase in pollen percentage of the tropical montane forest during the Middle Holocene, 5-4 ka BP, suggests a much cold condition along the Celebes Sea at the time. It is suggested that precipitation along the Celebes Sea has been affected by variations of land-ocean distribution and solar activities. It is implied by fluctuation in fern spores record that precipitation was at a high level during the Early Holocene, but lowered after the Middle Holocene, which is in a similar pattern as indicated by the Indonesian stalagmite records.
-
Key words:
- Holocene /
- Celebes Sea /
- pollen analysis /
- sea-level change /
- temperature /
- precipitation /
- climate change /
- vegetation
-
0. 引言
全新世是人类文明发展的重要时期,详细研究全新世的环境和气候变化,可为全球气候模拟、预测等提供不可或缺的资料.近20年来,大量对冰心、海洋、石笋、湖泊、黄土、珊瑚等记录的研究都聚焦于全新世的气候变化(Stott et al., 2002; Mayewski et al., 2004; Wang et al., 2005; Leduc et al., 2007; Partin et al., 2007; Griffiths et al., 2009; Wang and Li, 2009; Thevenon et al., 2010; Tierney et al., 2012).研究表明,早中全新世气候温暖湿润,与当前全球变暖下的环境特征类似,但全新世中期气候的不稳定性预示着未来全球气候系统有可能发生突变,这对全世界各国应对未来气候突变提出了新的挑战.同时,热带雨林区是全球重要的碳储库,其植被覆盖率高,生物多样性丰富,在热带以及全球气候系统中占有不可估量的作用(Maxwell and Liu, 2002; Bird et al., 2004; Hope et al., 2004; Anhuf et al., 2006; Bush et al., 2011).因此,作为植被研究重要手段之一的孢粉学研究也从高纬地区向中低纬度,从热带高山林地向低地植被延伸(Kershaw et al., 2007).海洋孢粉记录兼具陆地和海洋信息,并依托于海洋柱状样精细的沉积地层框架,可以体现晚第四纪,甚至更长时间尺度上的连续高分辨率的海陆环境和气候变化,不但可以用于重建古植被演化过程,还可以为海陆古气候和古环境记录的对比提供重要的依据(van der Kaars et al., 2000; Sun et al., 2003; Kershaw et al., 2007).
然而,目前低纬西太平洋地区全新世以来的植被变化主要集中在东南亚大陆地区和西太平洋岛屿,在海洋沉积中相对较少(Maxwell and Liu, 2002; Zhou et al., 2004; Wang et al., 2007; Lee and Liew, 2010; Xu et al., 2010).此外,已有的陆地孢粉记录连续性较差、海洋孢粉记录分辨率偏低,不足以用于揭示全新世时期完整清晰的植被演化及其对应的气候变化.如在加里曼丹、苏拉威西等湖泊沉积记录中,早中全新世的沉积记录不完整或者缺失,只能显示全新世晚期的植被、环境和气候变化(Anshari et al., 2001, 2004; Dam et al., 2001; Hope, 2001).本次工作对苏拉威西海周边地区全新世以来的海洋孢粉记录进行了深入的研究,通过区域性的海陆沉积记录对比,笔者得到了连续而详细的全新世植被演化以及相应的气候变化图景,为全新世快速气候变化的机制研究提供了新的证据.
1. 区域概况
位于苏拉威西海西部的婆罗洲属于热带高温湿润地区,适于大量的植被生长,拥有东南亚地区最为丰富的热带雨林植被,是地球上物种最多样的栖息地.在地形较为平坦的沿岸地区,潮水的影响范围可达内陆地区100 km.婆罗洲地区最高是海拔为4 102 m的基纳巴卢山.在婆罗洲低地地区年平均温度变化不大,但降雨量变化相对较为明显,而且存在分布差异.受西北季风影响,婆罗洲11月至次年4月的降雨量较大,稍微干旱时间段为6月至8月,但其月平均降雨量仍然在100 mm以上(MacKinnon et al., 1997).
由于该区域位于赤道季风气候及热带季风气候区内,热带雨林和季雨林是整个东南亚地区分布最广的2种植被类型,被称为“气候顶级群落”.热带地区植被面貌与温带明显不同,植被种类繁多,但每一种组合中没有明显的优势种,只有在一些特殊生境的植被类型及次生林中,才有少数的种类占据一定的优势(Hill, 1979).各地区生境条件的差异,对植被的生长和分布规律有一定的影响.因此,东南亚地区的植被可分为下列几种主要类型:热带雨林(rain forest)、热带季雨林(monsoon forest)、山地雨林(mountain vegetation of the rain forest)、山地季雨林(mountain vegetation of the monsoon forest)、红树林(mangrove)、砂质海滩林(beach forest)、硬叶常绿林(heath forest)、沼泽林(swamp forests)和次生林(secondary vegetations)(Hill, 1979; 严崇潮, 1984).其中热带雨林一般指地势海拔不超过1 200 m的植被,也被称为热带低地雨林(tropical lowland evergreen rain forest);山地雨林根据海拔分布可细分为海拔1 200~1 500 m的热带低山雨林(tropical lower montane rain forest)和海拔在1 500~3 000 m(或3 300 m)的热带高山雨林(tropical upper montane rain forest),以及超过3 000 m或3 350 m的热带亚高山森林(tropical subalpine forest)(Whitmore, 1984; 李逊和孙湘君,1999).
2. 研究材料及方法
本次研究选用的是1998年法国“Marion Dufresne”考察船MD111航次在苏拉威西海西北部采集的MD98-2178孔(3.6200°N,118.7000°E,水深1 984 m,总长35.6 m;图 1)样品.此次研究主要对深海钻孔的全新世样品进行孢粉分析和浮游有孔虫稳定氧同位素分析.孢粉分析的样品间距为20 cm.实验室处理过程为:首先用盐酸去除样品中的钙质部分,清洗后用氢氟酸浸泡样品以溶解硅质成分,再次清洗后用7 μm尼龙筛震荡过滤得到需鉴定的样品.为了计算孢粉的浓度, 在每个样品预处理前加入1粒外加石松药片.每个样品鉴定统计陆生种子植物花粉200粒以上,部分花粉较少的样品统计100粒左右,孢粉的百分含量计算是以陆生种子植物花粉总和为基数,其他各组分与该基数相比得到的百分含量.孢粉浓度是利用外加石松孢子来计算单位体积上的孢粉总量(grains/cm3).
 图 1 研究站位地理位置图中红星为站位位置,虚线为南海穿越流,实线为印尼穿越流;修改自Gordon et al.(2012)Fig. 1. General location of the study cores
图 1 研究站位地理位置图中红星为站位位置,虚线为南海穿越流,实线为印尼穿越流;修改自Gordon et al.(2012)Fig. 1. General location of the study cores浮游有孔虫氧同位素测定的实验过程包括:样品经过标准微体古生物学方法采集和预处理后,从大于63 μm的样品中,挑选出洁净无污染的壳径为300~360 μm的Globigerinoides ruber(G.ruber)壳体14枚.并在同济大学国家重点实验室中的无机质谱分析室内进行清洗和上机测试工作,有孔虫壳体在镜下用解剖针破碎后用乙醇在超声波浴中反复洗净,经40 ℃下干燥后在用MAT252质谱仪测定壳体中的δ18O值和δ13C值.分析精度采用中国国家标样GBW04405检测,氧、碳同位素标准偏差分别为±0.07‰,PDB和±0.04‰,PDB.
上述分析测试实验均在同济大学海洋地质国家重点实验室中完成.
3. 分析结果
3.1 年代框架
本次研究的年代框架是依据AMS14C测年结果.根据氧同位素记录,褚智慧(2011)在关键层位和有孔虫丰度较高的样品中挑选1 000枚壳径大于150 μm的G. ruber壳体,在德国基尔大学莱不尼茨(Leibniz)放射性实验室完成测试,共获得15个有效的AMS14C年龄值.根据AMS14C测年得到,MD98-2178孔上部700 cm代表了全新世的沉积记录(Fan et al., 2013;图 2).

3.2 孢粉组合类型
苏拉威西海西北部MD98-2178孔处于东南亚热带地区,周边陆地上的植物种类繁多,相应的,孢粉类型也很丰富.基于前人的植被考察和海陆孢粉研究资料(Hill, 1979; Whitmore, 1984; 严崇潮, 1984; Anshari et al., 2001, 2004)以及各植被的生态分布,本文将鉴定的花粉类型分为以下几个生态组合:
(1) 热带高山雨林(tropical upper montane rain forest).以叶枝杉(Phyllacladus)、罗汉松属(Podacarpus)和泪杉属(Dacrydium)为主,包括杨梅属(Myrica)、杜鹃属(Rhododendron)等.
(2) 热带低山雨林(tropical lower montane rain forest).以栎属(Quercus)、栲属(Castanopsis)和杜英科(Elaeocarpaceae)为主.
(3) 热带低地雨林(tropical lowland rain forest).以大戟科(Euphorbiaceae)、桑科(Moraceae)、棕榈科(Palmae)、木麻黄科(Casuarinaceae)、血桐属(Macaranga)、野桐属(Mallotus)、茜草科(Rubiaceae)、芸香科(Rutaceae)等为主;包括米仔兰属(Aglaia)、山麻杆属(Alchornea)、朴树属(Celtis)、旋花科(Convolvulac-eae)、五桠果属(Dillenia)、龙脑香科(Dipterocarpac-eae)、龙船花属(Ixora)、唇形科(Labiaceae)、无患子科(Sapindaceae)、楝科(Meliaceae)、桃金娘科(Myrtac-eae)、木犀科(Oleaceae)、椴树属(Tilia)、火把树科(Weinmannia)等.
(4) 红树林(mangrove).以红树属(Rhizoph-ora)为主,包括木榄属(Bruguiera)、海桑属(Sonneratia)等.
(5) 草本植被(grassland).以禾本科(Gramin-eae)和莎草科(Cyperaceae)为主,包括苋科(Amaranthaceae)、藜科(Chenopodiaceae)、菊科(Compasitae)、香蒲属(Typha)等.
在上述生态组合中,热带低地雨林的孢粉属种最多,但是热带低地雨林植被的花粉多数是虫媒传播,风媒传播的植被类型较少,导致各个科属的含量普遍偏低,变化幅度也较微弱.此外,科属间的花粉属性差异较大,在解释上存在很大的偏差.如东南亚低地雨林中占重要地位的龙脑香科(Dipterocarpaceae),在花粉图谱中的含量较低(图 3和图 4),解释时难免将其弱化.相对的桑科(Moraceae)、野桐属(Mallotus)、血桐属(Macaranga)、朴树属(Celtis)等花粉含量较高,在数据解释时会高估其相应的植被分布(Morley, 2002).
 图 3 苏拉威西海西北部MD98-2178孔孢粉百分比图谱除木榄属、红树属、叶枝杉、罗汉松属、栎属和桑科外, 其他科属孢粉含量刻度为0~ 10%Fig. 3. Pollen percentage records of core MD98-2178 from northwest Celebes Sea
图 3 苏拉威西海西北部MD98-2178孔孢粉百分比图谱除木榄属、红树属、叶枝杉、罗汉松属、栎属和桑科外, 其他科属孢粉含量刻度为0~ 10%Fig. 3. Pollen percentage records of core MD98-2178 from northwest Celebes Sea 图 4 苏拉威西海西北部MD98-2178孔孢粉浓度Fig. 4. Pollen concentration records of core MD98-2178 from northwest Celebes Sea
图 4 苏拉威西海西北部MD98-2178孔孢粉浓度Fig. 4. Pollen concentration records of core MD98-2178 from northwest Celebes Sea3.3 苏拉威西海西北部MD98-2178孔的全新世时期孢粉记录
该孔全新世的孢粉以红树林和热带低地雨林组合为主,蕨类孢子的含量也较高.自下而上,存在几个比较明显的变化过程.依据各属种花粉含量变化,以及Tilia分析软件中的CONISS分析结果,该花粉带可细分被为以下3个花粉亚带(图 3和图 4).
3.3.1 SU3带(深度700~420 cm,年龄为10.0~7.0 ka BP)
此亚带花粉记录显示,红树林花粉和蕨类孢子含量发生显著变化.红树林花粉的百分含量较高,在该花粉带初期为55%左右,最高可达62%,随后又小幅下降,在50%左右波动.其中以红树属花粉含量变化为主,同时木榄属也有明显变化.热带高山雨林的花粉含量在7%左右波动,以叶枝杉和罗汉松属花粉为主.热带低山雨林的花粉含量为10%左右,以栎属和栗属的花粉含量为主.热带低地雨林的花粉含量为20%,后期呈现上升趋势,科属以桑科和茜草科等为主.蕨类孢子含量持续增加,由初期的50%左右增加到后期的110%左右,而且仍然持续增长的趋势.从花粉浓度记录来看,除了蕨类外,其他各生态组合的花粉总量和花粉浓度是剧烈下降的,并且该趋势持续整个花粉带.蕨类孢子的浓度为350 grains/cm3(图 4).
3.3.2 SU2带(深度420~200 cm,年龄为7.0~4.4 ka BP)
花粉记录显示,该带以红树林和热带低地雨林为主.红树林花粉含量由55%下降到31% 左右,然后在该带后期花粉含量在35%附近波动,其中以红树属的花粉含量变化为主,同时木榄属也有明显变化.热带高山雨林的花粉含量由初期的11%上升到30%,叶枝杉和罗汉松属的花粉含量都有明显增加.草本花粉虽然有所增加,但其含量仍然低于5%.热带低地雨林中的大戟科、桑科等花粉含量有所上升.蕨类孢子的含量在初期时延续上一个花粉带的上升趋势,增加至180%,随后出现显著下降,降至150%.从孢粉浓度图谱来看,各生态组合的浓度都处于低值.
3.3.3 SU1带(深度200~10 cm,年龄为4.4~1.7 ka BP)
该带花粉含量与上个花粉带较为相似,只是红树林花粉含量有小幅上升,在41%左右波动.热带高山雨林花粉含量从上个花粉带末期的30%下降至10%左右.蕨类孢子含量在下降至70%后,回升至100%左右波动.从浓度变化来看,除热带高山雨林外,其他地区都表现为上升过程.
4. 讨论
从孢粉分析结果看,红树林、热带山地雨林、蕨类等植被类型的孢粉记录都发生了显著的变化,但各个孢粉组合在变化模式上存在明显差异.由此可见,各植被类型反馈了不同的气候条件或环境因子.下面对各孢粉组合所对应的植被演化和气候变化进行详细讨论,并将其与2006年中法合作的“Macro Polo Ⅱ”航次在菲律宾南部棉兰老岛Davao湾内采集的MD06-3075孔(6.476 2°N,125.832 2°E,水深1 878 m,总长30.76 m;图 1)全新世孢粉分析结果以及古海洋记录、石笋记录等进行对比(图 5和图 6),从而得到比较可靠的区域变化信息.
 图 5 苏拉威西海西北部MD98-2178孔与菲律宾南部MD06-3075孔海洋孢粉记录对比红色曲线以左侧坐标轴来度量;黑色曲线以右侧坐标轴来度量,据Bian et al.(2011);a.花粉总浓度;b.红树林花粉浓度;c.红树林花粉含量;d.热带高山雨林花粉含量;e.草本植被花粉含量;f.蕨类孢子含量;g.蕨类孢子浓度Fig. 5. Comparison of pollen records of core MD98-2178 from northwest Celebes Sea and MD06-3075 from the southern Philippines
图 5 苏拉威西海西北部MD98-2178孔与菲律宾南部MD06-3075孔海洋孢粉记录对比红色曲线以左侧坐标轴来度量;黑色曲线以右侧坐标轴来度量,据Bian et al.(2011);a.花粉总浓度;b.红树林花粉浓度;c.红树林花粉含量;d.热带高山雨林花粉含量;e.草本植被花粉含量;f.蕨类孢子含量;g.蕨类孢子浓度Fig. 5. Comparison of pollen records of core MD98-2178 from northwest Celebes Sea and MD06-3075 from the southern Philippines 图 6 MD98-2178孔海洋孢粉记录和古海洋记录与石笋记录对比黄色阴影区为全新世中期植被和气候突变期;a.浮游有孔虫氧同位素记录,据Fan et al.(2013);b.海水表层温度(SST),据Fan et al.(2013);c~e.热带高山雨林花粉含量、红树林花粉含量和蕨类孢子含量;f.蕨类孢子浓度;g.中国石笋δ18O记录,据Wang et al.(2005);h.婆罗洲石笋δ18O记录,据Partin et al.(2007);i.印度尼西亚石笋δ18O记录,据Griffiths et al.(2009)Fig. 6. Comparison of marine pollen and palaeoceanographic records from core MD98-2178 with stalagmite records
图 6 MD98-2178孔海洋孢粉记录和古海洋记录与石笋记录对比黄色阴影区为全新世中期植被和气候突变期;a.浮游有孔虫氧同位素记录,据Fan et al.(2013);b.海水表层温度(SST),据Fan et al.(2013);c~e.热带高山雨林花粉含量、红树林花粉含量和蕨类孢子含量;f.蕨类孢子浓度;g.中国石笋δ18O记录,据Wang et al.(2005);h.婆罗洲石笋δ18O记录,据Partin et al.(2007);i.印度尼西亚石笋δ18O记录,据Griffiths et al.(2009)Fig. 6. Comparison of marine pollen and palaeoceanographic records from core MD98-2178 with stalagmite records4.1 离岸距离和源区植被结构的差异
由于两个站位的植被类型比较相似,因此孢粉总体传播能力是相似的.但是从两个站位的孢粉记录对比来看,MD98-2178孔的红树林含量高于MD06-3075孔,但MD06-3075孔的蕨类孢子含量和浓度却远远高于MD98-2178孔.比如,菲律宾南部MD06-3075孔蕨类孢子的含量最高可达700%,浓度在2 000~3 000 grains/cm3波动(Bian et al., 2011;图 5).相对苏拉威西海西北部MD98-2178孔蕨类孢子含量最高值仅为270%,浓度高值只有500 grains/cm3左右.
上述孢粉浓度值差异与两个站位的离岸距离有很大关系.文献显示,MD06-3075孔离岸距离约30 km,并且湾内没有明显的陆架区域,而MD98-2178孔离岸距离为100~200 km,其中陆架地区延伸了大约50 km(边叶萍等, 2012).从海洋表层样分析来看,热带孢粉浓度高值基本在离岸较近的区域,并且离岸越远,浓度下降明显(van der Kaars and De Deckker, 2003).因此,两个站位与各自植被源区之间的距离不同,导致孢粉浓度上存在巨大的差距.
但是孢粉的含量变化差异,与源区各植被结构有很大关系.比如,红树林MD98-2178孔的红树林含量远远高于MD06-3075孔,而且在全新世早期,MD98-2178孔的红树林浓度甚至高于MD06-3075孔(图 5b和5c).这只能说明在苏拉威西海西北部地区,即婆罗洲东部沿岸,红树林的分布范围要远远超过了菲律宾南部沿岸地区.事实上,从现代红树林分布来看,婆罗洲东部沿岸特别是在几个河口三角洲地区,其红树林延伸范围很广;而相对在菲律宾南部Davao湾内,红树林分布范围则比较局限(Grindrod et al., 2002).因此,从各孢粉组合含量记录来看,苏拉威西海西北地区相对于菲律宾南部,其植被以红树林为主,热带山地雨林和热带雨林植被分布也较广,但草本植被和蕨类植被则相对较少.
4.2 全新世时期的海平面变化
红树林花粉记录显示,在全新世早期,其花粉含量较高(图 3和图 5c),体现了该植被类型相对于其他植被类型,其分布范围较广.这主要是由于延续冰消期海平面上升过程中,内陆地区受海水影响的地区扩大,适合红树林生长的空间增加造成的,但红树林花粉浓度却表现为急剧下降(图 4和图 5b),其可能体现了海平面变化对沉积距离的影响.到了大约7 ka BP,花粉的含量出现明显下降,同时其浓度达到低谷(图 3和图 4).而MD06-3075孔的红树林花粉含量从全新世早期一直到6 ka BP都较高,红树林的分布范围很广,也较为稳定;在6.0~4.5 ka BP期间,花粉含量下降可能说明了红树林发展明显受到限制;此外,MD06-3075孔的红树林花粉浓度变化模式与花粉含量的变化模式相似(Bian et al., 2011;图 5b和5c).同时,比较两个站位各孢粉浓度可知,苏拉威西海西北部MD98-2178孔各孢粉组合的浓度变化过程是类似的,都是先下降,稳定一定时间后再上升,而MD06-3075孔的各孢粉组合间的浓度变化差异大.
上述这些差异很可能是由海平面变化以及两个站位所处地形差异所造成的(边叶萍等, 2012).由于婆罗洲东部的地势较为平坦,陆架较菲律宾南部宽广,在海平面上升期间,陆架被逐渐淹没,海岸线逐渐退向内陆地区,导致红树林的生长空间也向内陆迁移,与沉积点的距离加大,从而使海洋孢粉记录中的红树林花粉含量和浓度明显下降.但是,在海平面影响不大的情况下,海洋孢粉中红树林花粉记录可能更多地反映气候变化.因此,MD98-2178孔的红树林变化受海平面影响较大,而MD06-3075孔红树林记录可能反映了气候因子,如降雨量变化(Bian et al., 2011).
由此可见,MD98-2178孔的红树林花粉含量和孢粉浓度记录显示了该地区的海平面变化:全新世早期至7 ka BP时期,孢粉浓度下降,以及红树林花粉含量的高值,可能指示海平面在此阶段一直处于上升过程;而在7~4 ka BP期间,浓度低值则说明该时期为相对高海平面时期;在4 ka BP之后,苏拉威西海西北部的红树林花粉含量和浓度都有微弱的上升,可能指示海平面的小幅下降.
4.3 全新世中期的降温事件
热带高山雨林位于较高海拔地区,是距离沉积地点较远的类群,但叶枝杉(Phyllacladus)、罗汉松属(Podacarpus)等花粉都属于气囊型花粉,主要以风媒传播为主,传播距离相对其他类型的花粉要远.同时,气囊型花粉搬运过程中受其他气候因素的影响也较小.根据其生态特征,当气候变冷时,它的植被带向下迁移,以适应低海拔地区较暖的、与其所适应的环境更相似的温度条件.这时高山雨林花粉的传播路径变短,而且低海拔区的适宜面积也增大,传播到沉积地点的该类花粉的含量增高.反之,当气候变暖时其生长范围上移,这类花粉含量减少.虽然其他花粉类型也有类似的变化,但由于这种变化对于热带高山雨林表现得更明显,所以其花粉含量变化指示了植被带的迁移和温度变化.
孢粉结果显示,苏拉威西海西北部MD98-2178孔热带高山雨林的花粉含量在6 ka BP左右开始呈现上升趋势,特别是在5~4 ka BP时期,含量高达30%,明显高于全新世早期(5%~10%),说明此时热带高山雨林的分布范围向低海拔地区扩张,体现站位周边地区温度有所下降(图 5和图 6c).尽管MD06-3075孔热带高山雨林花粉含量在6~5 ka BP时期呈现一个峰值,但变化幅度明显小于MD98-2178孔.从两个地区全新世海水表层温度记录来看,菲律宾南部的海水表层温度变化是逐渐下降的,但变化幅度不大(Stott et al., 2004).而苏拉威西海西北部的海水表层温度在全新世中期下降了1 ℃左右(Fan et al., 2013;图 6b).考虑到菲律宾南部位于西太平洋暖池的中心地带,而苏拉威西海西北部的MD98-2178孔则处于暖池的边缘地区,对于温度变化会相对比较敏感.因此,从整体上来看,海洋孢粉也记录了全新世中期时,即5~4 ka BP,该区域存在一个温度低值期.
4.4 苏拉威西海周边地区的降雨量变化
学者对于全新世时期西太平洋热带地区的降雨量变化一直存在争议.主要是由于该地区常年降雨量较高,变化幅度相对于高纬地区要小.同时,该地区受到东亚季风、印度季风、澳洲季风、ENSO(El Niño Southern Oscillation)等气候系统的影响,降雨量的控制因素显得尤为复杂(Steinke et al., 2014).比如,东南亚地区全新世时期的夏季降雨量与太阳辐射量变化关系密切;澳大利亚-印度尼西亚地区的降雨量变化则受到太阳辐射和海洋反馈(主要是海陆分布格局和海水表层温度等变化)的共同作用(Liu et al., 2004).这在中国石笋和印尼石笋等记录可以有十分明显的体现(Wang et al., 2005; Griffiths et al., 2009;图 6g和6i).但是,位于亚洲季风和澳洲季风交汇处的婆罗洲石笋记录却与上述石笋记录都不同.对此,笔者希望利用其他的降雨量指标来探讨该地区的降雨量变化,为苏拉威西海周边地区降雨量变化的控制因素提供更多的证据.事实上,热带植被对于降雨量的变化也是十分敏感的,草本植被、蕨类孢子等变化一直都用于指示植被源区的降雨量变化.本次研究中两个站位完整高分辨率的海洋孢粉记录可提供该地区的降雨量变化信息.
菲律宾南部MD06-3075孔的孢粉记录显示,在全新世早期气候呈现暖湿状态.但在全新世中期,红树林退缩、蕨类含量下降以及草本增加等植被变化(图 5),这些可能反映了该地区河流径流量的减少,在一定程度上证实了影响菲律宾南部地区降雨的东亚季风减弱和ENSO系统增强导致降雨量下降的论点(Bian et al., 2011).
但是,由于苏拉威西海西北部MD98-2178孔全新世时期的红树林花粉记录受海平面影响较大,而且其草本植被的含量非常低(基本低于5%),所以该站位可反映降雨量变化的主要是蕨类孢子记录.在10~6 ka BP时期,MD98-2178孔蕨类孢子含量从30%增加至270%左右,而浓度从400 grains/cm3下降至200 grains/cm3(图 6e和6f).尽管其浓度是下降的,但是比同时期花粉总浓度变化幅度要小(图 5a和5g).因此,从总体上来看,在全新世早期蕨类分布是持续增加的,体现了全新世早期温暖湿润的气候状态.而在6~4 ka BP时期,蕨类孢子含量的下降说明其分布范围有所减少,体现了该地区湿度有所下降.
另一方面,尽管陆地孢粉记录的连续性较差,但是从该地区的陆地研究结果来看,在加里曼丹和苏拉威西岛等湖泊沉积和孢粉沉积记录中,大约5 ka BP以来炭屑含量和湖平面则分别表现为上升和下降过程(Dam et al., 2001; Hope, 2001; Anshari et al., 2001, 2004).同时,从GPCC观测的季节性降雨量变化来看,菲律宾南部地区北半球夏季降雨量较高,印尼地区的北半球冬季降雨量较高.婆罗洲地区尽管季节差异较小,但是相对干季是在北半球冬季时期(图 7).国际古气候模拟比较计划(paleoclimate model intercomparison project, PMIP)中的11个海气耦合模式的模拟结果显示,菲律宾南部的北半球夏季(6月~8月)降雨量在全新世中期是有所降低的.婆罗洲地区的季节性降雨量变化显示,全新世中期时,北半球夏季(6月~8月)时的降雨量增加,但是北半球冬季(12月~次年2月)时降雨量是下降的(Tierney et al., 2012).因此,陆地孢粉记录和数值模拟与MD98-2178孔的海洋孢粉记录都指示了全新世中期苏拉威西海西部地区的降雨量是下降的.
 图 7 菲律宾南部棉兰老岛(a)、婆罗洲(b)以及印度尼西亚Flores岛的月平均降雨量变化(c)降雨量资料来源于全球降水气候中心(Global Precipitation Climatology Centre,GPCC,http://gpcc.dwd.de);据Tierney et al.(2012)资料Fig. 7. Observed monthly cycles of precipitation for the Mindanao in the southern Philippines (a), Borneo (b), and Flores in Indonesia (c)
图 7 菲律宾南部棉兰老岛(a)、婆罗洲(b)以及印度尼西亚Flores岛的月平均降雨量变化(c)降雨量资料来源于全球降水气候中心(Global Precipitation Climatology Centre,GPCC,http://gpcc.dwd.de);据Tierney et al.(2012)资料Fig. 7. Observed monthly cycles of precipitation for the Mindanao in the southern Philippines (a), Borneo (b), and Flores in Indonesia (c)综上所述,尽管全新世早期苏拉威西海周边地区的降雨量是持续增加的,但在全新世中期,即6~4 ka BP时期,海陆孢粉记录和数值模拟结果都显示降雨量的减少.这种变化过程表明,菲律宾南部MD06-3075孔附近的降雨量变化受东亚季风的影响较大;而苏拉威西海西北部MD98-2178孔附近地区的降雨量变化模式与印尼石笋记录较为相似,其全新世时期的变化与婆罗洲石笋记录不同,说明该地区降雨量受全新世早期海陆格局和全新世中期太阳活动影响较大.由于陆地孢粉记录和数值模拟结果与海洋孢粉记录一致,因此对于婆罗洲石笋记录所反映的降雨量变化过程及其控制因素,可能还需要更多的区域性资料进行综合和进一步的研究.
5. 结论
本次研究利用MD98-2178孔高分辨率的海洋孢粉和稳定氧同位素记录,重建了苏拉威西海周边地区全新世时期的热带植被和气候环境演变,通过海陆沉积记录对比,得到以下几点认识:
孢粉分析结果显示,红树林花粉含量反映了地区间红树林分布范围的不同.MD98-2178孔较高含量的红树林花粉表明婆罗洲东部红树林较为繁盛,这与陆架较菲律宾南部宽广有关.同时,这种地形特征导致了苏拉威西海MD98-2178孔的红树林花粉记录比菲律宾南部MD06-3075孔更易受到海平面变化影响.孢粉记录显示全新世早期至7 ka BP时期,海平面处于上升阶段,在7~4 ka BP为高海平面期,在4 ka BP之后,海平面有小幅的下降.
热带高山雨林花粉记录显示,在全新世中期,即5~4 ka BP,苏拉威西海西部地区存在一个温度低值期.由于站位与西太平洋暖池中心区的相对位置,该地区温度下降幅度较为显著.
关于苏拉威西海周边地区的降雨量变化,MD98-2178孔蕨类孢子含量变化显示苏拉威西海西部地区降雨量在全新世早期是持续增加的,但在全新世中期,即6~4 ka BP时期,降雨量有所减少,体现了全新世该地区降雨量受海陆格局和太阳活动共同影响.而菲律宾南部MD06-3075孔附近的降雨量变化受东亚季风的影响较大.
致谢: 本次研究中孢粉鉴定和分析工作得到了同济大学海洋地质国家重点实验室孙湘君教授、翁成郁教授、澳大利亚国立大学太平洋和亚洲研究院Janelle Stevenson教授等的精心指导和无私帮助,在此表示衷心感谢!同时也要感谢对本文提供宝贵意见和建议的评审专家们和编委们. -
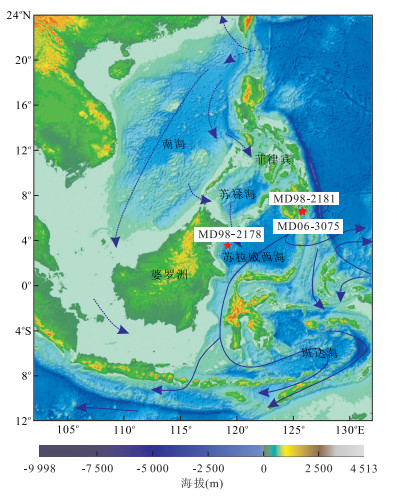
图 1 研究站位地理位置
图中红星为站位位置,虚线为南海穿越流,实线为印尼穿越流;修改自Gordon et al.(2012)
Fig. 1. General location of the study cores

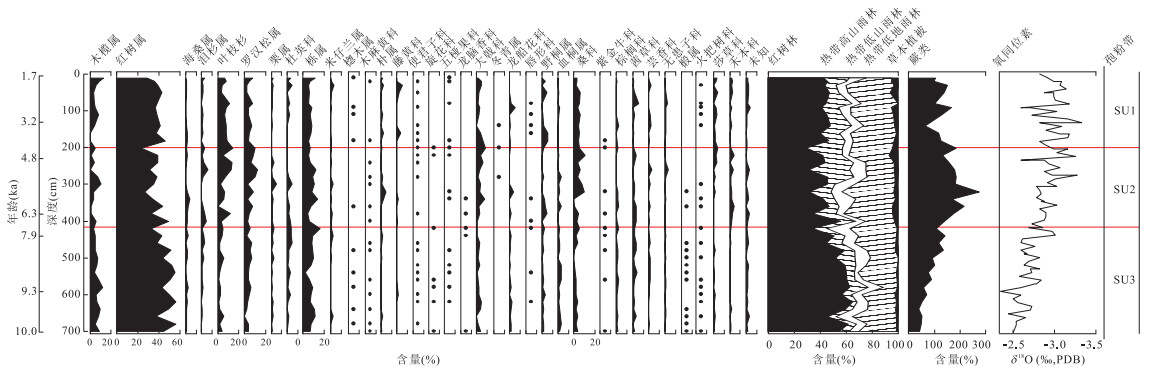
图 3 苏拉威西海西北部MD98-2178孔孢粉百分比图谱
除木榄属、红树属、叶枝杉、罗汉松属、栎属和桑科外, 其他科属孢粉含量刻度为0~ 10%
Fig. 3. Pollen percentage records of core MD98-2178 from northwest Celebes Sea

图 4 苏拉威西海西北部MD98-2178孔孢粉浓度
Fig. 4. Pollen concentration records of core MD98-2178 from northwest Celebes Sea
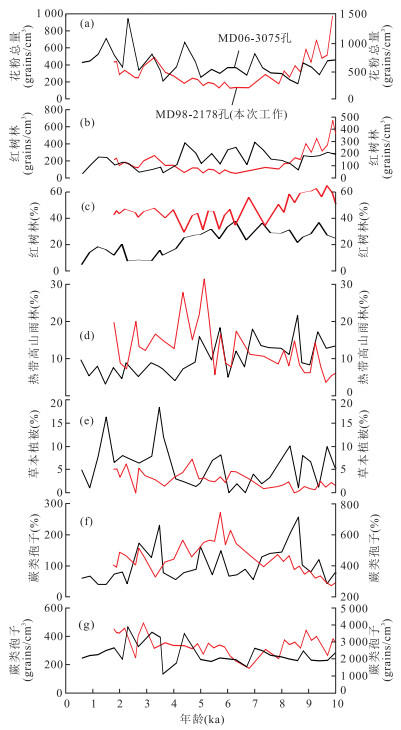
图 5 苏拉威西海西北部MD98-2178孔与菲律宾南部MD06-3075孔海洋孢粉记录对比
红色曲线以左侧坐标轴来度量;黑色曲线以右侧坐标轴来度量,据Bian et al.(2011);a.花粉总浓度;b.红树林花粉浓度;c.红树林花粉含量;d.热带高山雨林花粉含量;e.草本植被花粉含量;f.蕨类孢子含量;g.蕨类孢子浓度
Fig. 5. Comparison of pollen records of core MD98-2178 from northwest Celebes Sea and MD06-3075 from the southern Philippines
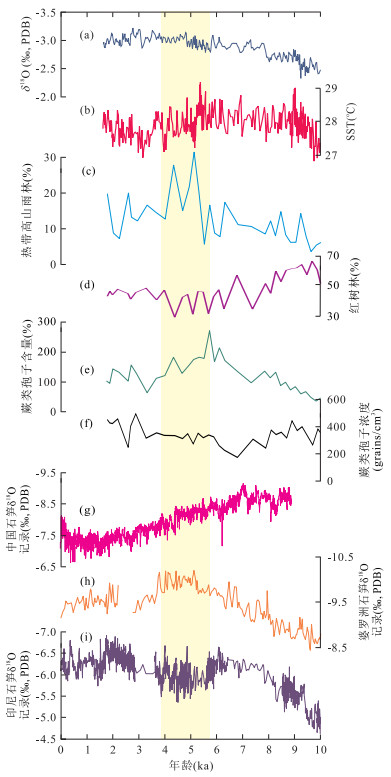
图 6 MD98-2178孔海洋孢粉记录和古海洋记录与石笋记录对比
黄色阴影区为全新世中期植被和气候突变期;a.浮游有孔虫氧同位素记录,据Fan et al.(2013);b.海水表层温度(SST),据Fan et al.(2013);c~e.热带高山雨林花粉含量、红树林花粉含量和蕨类孢子含量;f.蕨类孢子浓度;g.中国石笋δ18O记录,据Wang et al.(2005);h.婆罗洲石笋δ18O记录,据Partin et al.(2007);i.印度尼西亚石笋δ18O记录,据Griffiths et al.(2009)
Fig. 6. Comparison of marine pollen and palaeoceanographic records from core MD98-2178 with stalagmite records
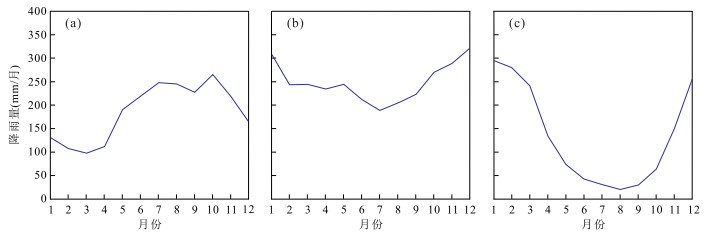
图 7 菲律宾南部棉兰老岛(a)、婆罗洲(b)以及印度尼西亚Flores岛的月平均降雨量变化(c)
降雨量资料来源于全球降水气候中心(Global Precipitation Climatology Centre,GPCC,http://gpcc.dwd.de);据Tierney et al.(2012)资料
Fig. 7. Observed monthly cycles of precipitation for the Mindanao in the southern Philippines (a), Borneo (b), and Flores in Indonesia (c)
-
Anhuf, D., Ledru, M.P., Behling, H., et al., 2006. Paleo-Environmental Change in Amazonian and African Rainforest during the LGM. Palaeogeography, Palaeoclimatology, Palaeoecology, 239(3-4): 510-527. doi: 10.1016/j.palaeo.2006.01.017 Anshari, G., Kershaw, A.P., van der Kaars, S., 2001. A Late Pleistocene and Holocene Pollen and Charcoal Record from Peat Swamp Forest, Lake Sentarum Wildlife Reserve, West Kalimantan, Indonesia. Palaeogeography, Palaeoclimatology, Palaeoecology, 171(3-4): 213-228. doi: 10.1016/S0031-0182(01)00246-2 Anshari, G., Kershaw, A.P., van der Kaars, S., et al., 2004. Environmental Change and Peatland Forest Dynamics in the Lake Sentarum Area, West Kalimantan, Indonesia. Journal of Quaternary Science, 19(7): 637-655. doi: 10.1002/jqs.879 Bian, Y.P., Jian, Z.M., Weng, C.Y., et al., 2011. A Palynological and Palaeoclimatological Record from the Southern Philippines since the Last Glacial Maximum. Chinese Science Bulletin, 56(22): 2359-2365. doi: 10.1007/s11434-011-4573-1 Bian, Y.P., Li, J.B., Jian, Z.M., et al., 2012. The Different Response of Marine Pollen Records to the Sea Level Change in the Low Latitude West Pacific since the Last Glacial Period. Quaternary Sciences, 32(6): 1078-1086 (in Chinese with English abstract). http://en.cnki.com.cn/Article_en/CJFDTOTAL-DSJJ201206005.htm Bird, M.I., Hope, G., Taylor, D., 2004. Polulating PEP Ⅱ: The Dispersal of Humans and Agriculture through Austral-Asia and Oceania. Quaternary International, 118-119: 145-163. doi: 10.1016/S1040-6182(03)00135-6 Bush, M., Flenley, J.R., Gosling, W., 2011. Tropical Rainforest Responses to Climatic Change. Praxis Publishing, Chichester. Chu, Z.H., 2011. Late Quaternary Responses of Upper Ocean Water to the Global Climate Changes in the Sulawesi Sea (Dissertation). Tongji University, Shanghai (in Chinese with English abstract). Dam, R.A.C., Fluin, J., Suparan, P., et al., 2001. Palaeoenvironmental Developments in the Lake Tondano Area (N. Sulawesi, Indonesia) since 33 000 B.P. . Palaeogeography, Palaeoclimatology, Palaeoecology, 171(3-4): 147-183. doi: 10.1016/S0031-0182(01)00244-9 Fan, W.J., Jian, Z.M., Bassinot, F., et al., 2013. Holocene Centennial-Scale Changes of the Indonesian and South China Sea Throughflows: Evidences from the Makassar Strait. Global and Planetary Change, 111: 111-117. doi: 10.1016/j.gloplacha.2013.08.017 Grindrod, J., Moss, P., van der Kaars, S., 2002. Mangrove Palynology in Continental Shelf and Deep Sea Cores of the North Australian-Indonesian Region. In: Kershaw, P., David, B., Tapper, N., et al., eds., Bridging Wallace's Line: The Environmental and Cultural History and Dynamics of the SE-Asian-Australian Region. Catena-Verlag, Reiskirchen. Griffiths, M.L., Drysdale, R.N., Gagan, M.K., et al., 2009. Increasing Australian-Indonesian Monsoon Rainfall Linked to Early Holocene Sea-Level Rise. Nature Geoscience, 2(9): 636-639. doi: 10.1038/ngeo605 Gordon,A.L.,Huber,B.A.,Metzger,E.J.,et al.,2012.South China Sea Throughflow Impact on the Indonesian Throughflow.Geophysical Research Letters,39:L11602.doi: 10.1029/2012GL052021.doi:10.1029/2012GL052021,2012 Hill, R.D., 1979. South-East Asia: A Systematic Geography. Oxford University Press, Oxford. Hope, G., 2001. Environmental Change in the Late Pleistocene and Later Holocene at Wanda Site, Soroako, South Sulawesi, Indonesia. Palaeogeography, Palaeoclimatology, Palaeoecology, 171(3-4): 129-145. doi: 10.1016/S0031-0182(01)00243-7 Hope, G., Kershaw, A.P., van der Kaars, S., et al., 2004. History of Vegetation and Habitat Change in the Austral-Asian Region. Quaternary International, 118-119: 103-126. doi: 10.1016/S1040-6182(03)00133-2 Kershaw, A.P., van der Kaars, S., Flenley, J.R., 2007. The Quaternary History of Far Eastern Rainforests. In: Bush, M.B., Flenley, J.R., eds., Tropical Rainforest Responses to Climatic Change. Praxis Publishing, Chichester, 77-115. Leduc, G., Vidal, L., Tachikawa, K., et al., 2007. Moisture Transport across Central America as a Positive Feedback on Abrupt Climatic Changes. Nature, 445(7130): 908-910. doi: 10.1038/nature05578 Lee, C.Y., Liew, P.M., 2010. Late Quaternary Vegetation and Climate Changes Inferred from a Pollen Record of Dongyuan Lake in Southern Taiwan. Palaeogeography, Palaeoclimatology, Palaeoecology, 287(1-4): 58-66. doi: 10.1016/j.palaeo.2010.01.015 Li, X., Sun, X.J., 1999. Palynological Records since Last Glacial Maximum from a Deep Sea Core in Southern South China Sea. Quaternary Sciences, (6): 526-535(in Chinese with English abstract). http://www.oalib.com/paper/1571012 Liu, Z., Harrison, S.P., Kutzbach, J., et al., 2004. Global Monsoons in the Mid-Holocene and Oceanic Feedback. Climate Dynamics, 22(2-3): 157-182. doi: 10.1007/s00382-003-0372-y MacKinnon, K., Hatta, G., Halim, H., et al., 1997. The Ecology of Kalimantan, Indonesia Borneo. Oxford University Press, Oxford. Maxwell, A.L., Liu, K.B., 2002. Late Quaternary Pollen and Associated Records from the Monsoonal Areas of Continental South and SE Asia. In: Kershaw, P., David, B., Tapper, N., et al., eds., Bridging Wallace's Line: The Environmental and Cultural History and Dynamics of the SE-Asian-Australian Region. Catena-Verlag, Reiskirchen. Mayewski, P.A., Rohling, E.E., Stager, J.C., et al., 2004. Holocene Climate Variability. Quaternary Research, 62(3): 243-255. doi: 10.1016/j.yqres.2004.07.001 Morley, R.J., 2002. Tertiary Vegetation History of Southeast Asia, with Emphasis on the Biogeographical Relationships with Australia. In: Kershaw, P., David, B., Tapper, N., et al., eds., Bridging Wallace's Line: The Environmental and Cultural History and Dynamics of the SE-Asian-Australian Region. Catena Verlag, Reiskirchen. Partin, J.W., Cobb, K.M., Adkins, J.F., et al., 2007. Millennial-Scale Trends in West Pacific Warm Pool Hydrology since the Last Glacial Maximum. Nature, 449(7161): 452-456. doi: 10.1038/nature06164 Steinke, S., Mohtadi, M., Prange, M., et al., 2014. Mid- to Late-Holocene Australian-Indonesian Summer Monsoon Variability. Quaternary Science Reviews, 93: 142-154. doi: 10.1016/j.quascirev.2014.04.006 Stott, L., Cannariato, K., Thunell, R., et al., 2004. Decline of Surface Temperature and Salinity in the Western Tropical Pacific Ocean in the Holocene Epoch. Nature, 431(7004): 56-59. doi: 10.1038/nature02903 Stott, L., Poulsen, C., Lund, S., et al., 2002. Super ENSO and Global Climate Oscillations at Millennial Time Scales. Science, 297(5579): 222-226. doi: 10.1126/science.1071627 Sun, X.J., Luo, Y.L., Huang, F., et al., 2003. Deep-Sea Pollen from the South China Sea: Pleistocene Indicators of East Asian Monsoon. Marine Geology, 201(1-3): 97-118. doi: 10.1016/S0025-3227(03)00211-1 Thevenon, F., Williamson, D., Bard, E., et al., 2010. Combining Charcoal and Elemental Black Carbon Analysis in Sedimentary Archives: Implications for Past Fire Regimes, the Pyrogenic Carbon Cycle, and the Human-Climate Interactions. Global and Planetary Change, 72(4): 381-389. doi: 10.1016/j.gloplacha.2010.01.014 Tierney,J.E.,Oppo,D.W.,LeGrande,A.N.,et al.,2012.The Influence of Indian Ocean Atmospheric Circulation on Warm Pool Hydroclimate during the Holocene Epoch.Journal of Geophysical Research,117(D19):D19108.doi: 10.1029/2012JD018060.doi:10.1029/2012JD018060 van der Kaars, S., De Deckker, P., 2003. Pollen Distribution in Marine Surface Sediments Offshore Western Australia. Review of Palaeobotany and Palynology, 124(1-2): 113-129. doi: 10.1016/s0034-6667(02)00250-6 van der Kaars, S., Wang, X., Kershaw, P., et al., 2000. A Late Quaternary Palaeoecological Record from the Banda Sea, Indonesia: Patterns of Vegetation, Climate and Biomass Burning in Indonesia and Northern Australia. Palaeogeography, Palaeoclimatology, Palaeoecology, 155(1-2): 135-143, 147-153. doi: 10.1016/S0031-0182(99)00098-X Wang, P.X., Li, Q.Y., 2009. The South China Sea, Paleoceanography and Sedimentology. Springer, Dordrecht. Wang, S.Y., Lü, H.Y., Liu, J.Q., 2007. The Early Holocene Optimum Inferred from a High-Resolution Pollen Record of Huguanyan Maar Lake in Southern China. Chinese Science Bulletin, 52(20): 2829-2836. doi: 10.1007/s11434-007-0419-2 Wang, Y.J., Cheng, H., Edwards, R.L., et al., 2005. The Holocene Asian Monsoon: Links to Solar Changes and North Atlantic Climate. Science, 308(5723): 854-857. doi: 10.1126/science.1106296 Whitmore, T.C., 1984. Tropical Rain Forests of the Far East. Clarendon Press, Oxford, 121-163. Xu, D.K., Lu, H.Y., Wu, N.Q., et al., 2010.30 000-Year Vegetation and Climate Change around the East China Sea Shelf Inferred from a High-Resolution Pollen Record. Quaternary International, 227(1): 53-60. doi: 10.1016/j.quaint.2010.04.015 Yan, C.C., 1984. Types and Distribution Patterns of Vegetation in Southeastern Asia. Chinese Journal of Ecology, (5): 28-32(in Chinese with English abstract). http://en.cnki.com.cn/Article_en/CJFDTotal-STXZ198405006.htm Zhou, W.J., Yu, X.F., Jull, A.J.T., et al., 2004. High-Resolution Evidence from Southern China of an Early Holocene Optimum and a Mid-Holocene Dry Event during the Past 18 000 Years. Quaternary Research, 62(1): 39-48. doi: 10.1016/j.yqres.2004.05.004 边叶萍, 李家彪, 翦知湣, 等, 2012. 低纬西太平洋末次冰期以来海洋孢粉记录对海平面变化的不同响应. 第四纪研究, 32(6): 1078-1086. doi: 10.3969/j.issn.1001-7410.2012.06.03 褚智慧, 2011. 苏拉威西海晚第四纪上层海水对全球气候变化的响应(硕士学位论文). 上海: 同济大学. 李逊, 孙湘君, 1999. 南海南部末次冰期以来的孢粉记录及其气候意义. 第四纪研究, 6: 526-535. doi: 10.3321/j.issn:1001-7410.1999.06.005 严崇潮, 1984. 东南亚的植被类型和分布规律. 生态学杂志, (5): 28-32. https://www.cnki.com.cn/Article/CJFDTOTAL-STXZ198405006.htm 期刊类型引用(1)
1. 李文宝,王汝建. 近2 Ma BP以来地球轨道参数周期上全球海平面变化机制. 地球科学. 2016(05): 742-756 .  本站查看
本站查看其他类型引用(2)
-

 下载:
下载:






 下载:
下载:
 点击查看大图
点击查看大图
计量
- 文章访问数: 4143
- HTML全文浏览量: 621
- PDF下载量: 432
- 被引次数: 3
