






The Biostratigraphic Correlation of the Permian-Triassic Boundary in Jinzhong Section, Weining, Guizhou, South China
-
摘要: 二叠纪—三叠纪之交发生了显生宙最大的生物灭绝事件,同时发生了复杂的环境变化.由于陆相二叠系-三叠系界线附近生物化石记录没有海相记录完整,同时也缺乏其他可靠的年代地层标志,所以如何标定陆相剖面的二叠纪—三叠纪界线一直是研究热点和难点.贵州威宁金钟地区二叠纪—三叠纪之交位于康滇古陆的东侧,毗邻海陆过渡相区,可以与同期的浅海碎屑岩相、海陆过渡相、陆相剖面进行由海到陆“追索式”的生物地层对比,是我国研究海、陆相二叠系-三叠系界线及对比的关键研究区之一.研究区的金钟二叠系-三叠系界线剖面中化石丰富,共发现双壳类化石4属6种,叶肢介2属4种,植物化石9属及腕足类化石1属,为海、陆相地层对比提供了桥梁.依据叶肢介Euestheria gutta-Palaeolimnadia xuanweiensis组合、植物Annalepis-Peltaspermum组合以及双壳类Pteria ussurica variabilis-Neoschizodus orbicularis组合的生物化石时代归属讨论,识别出金钟剖面的二叠系-三叠系界线应该在卡以头组下部.通过与邻区不同相剖面的对比,提出双壳类Pteria ussurica variabilis-Neoschizodus orbicularis组合以及陆相的叶肢介Euestheria gutta-Palaeolimnadia xuanweiensis组合可以作为三叠纪最早期的识别和对比标志.Abstract: The Permian-Triassic extinction event, which was associated with major global environmental changes, was the most significant one in the Phanerozoic. However, fossil records around the Permian-Triassic boundary in the terrestrial section are not complete compared with those in the marine section, and the precise time framework is not available. Thus the study of the Permian-Triassic boundary in terrestrial section has been hot and complicated. The Jinzhong section located in the eastern margin of the Kangdian Continent, near the transitional facies zone of the Permian and Triassic, can be compared with the marine section and terrestrial section at the same time in this period. The Jinzhong section has abundant fossils, including bivalve (4 genus 6 species), conchostracans (2 genus 4 species), plant fossils (9 genus) and brachiopods (1 genus).According to the age of the conchostracans fossil assemblages Euestheria gutta-Palaeolimnadia xuanweiensis, the plant fossil assemblages Annalepis sp.-Peltaspermum sp. and the bivalve fossil assemblages Pteria ussurica variabilis-Neoschizodus orbicularis, the Permian-Triassic boundary in the Jinzhong section is at the bottom of Kayitou Formation. In addition, the conchostracans fossil assemblages Euestheria gutta-Palaeolimnadia xuanweiensis and the bivalve fossil assemblages Pteria ussurica variabilis-Neoschizodus orbicularis can be used as a reliable marker of early Triassic in stratigraphic division and correlation in the marine-continental transitional facies in South China.
-
Key words:
- Permian-Triassic boundary /
- biostratigraphy /
- paleobiolgy /
- Jinzhong section /
- stratigraphy /
-
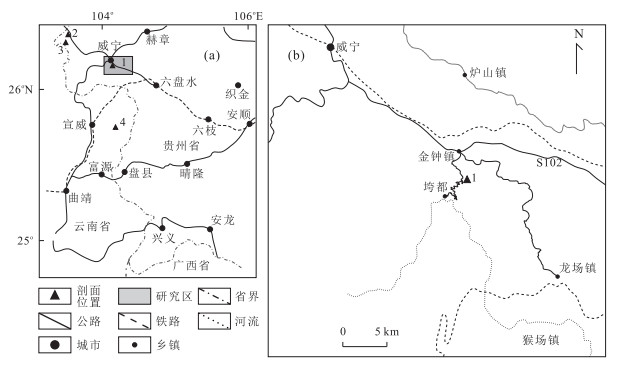
图 1 贵州威宁金钟剖面地理位置
a.研究区位置;b.研究剖面位置;1.金钟剖面;2.小河边剖面;3.岔河剖面;4.密得剖面
Fig. 1. Geographical position of Jinzhong Section, Weining, Guizhou
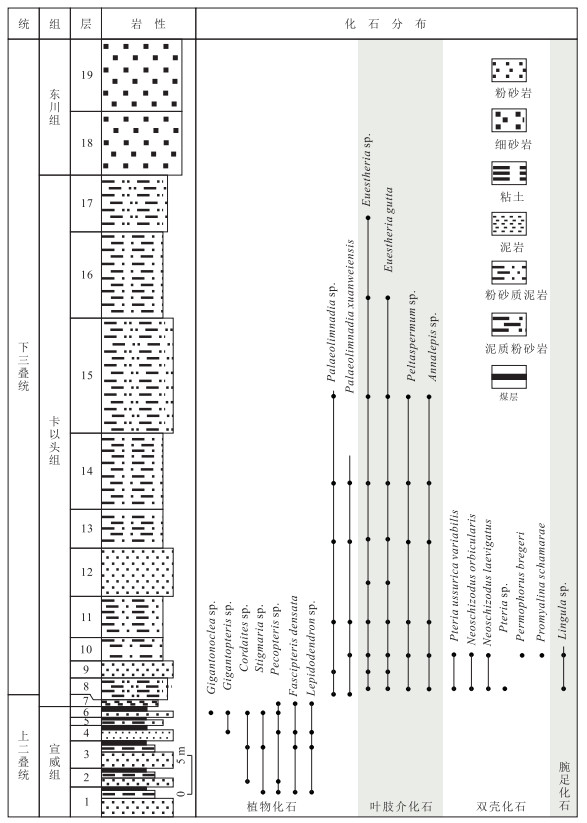
图 2 贵州威宁金钟剖面岩性柱状图及化石分布
Fig. 2. Lithology and biostratigraphy of Jinzhong Section, Weining, Guizhou
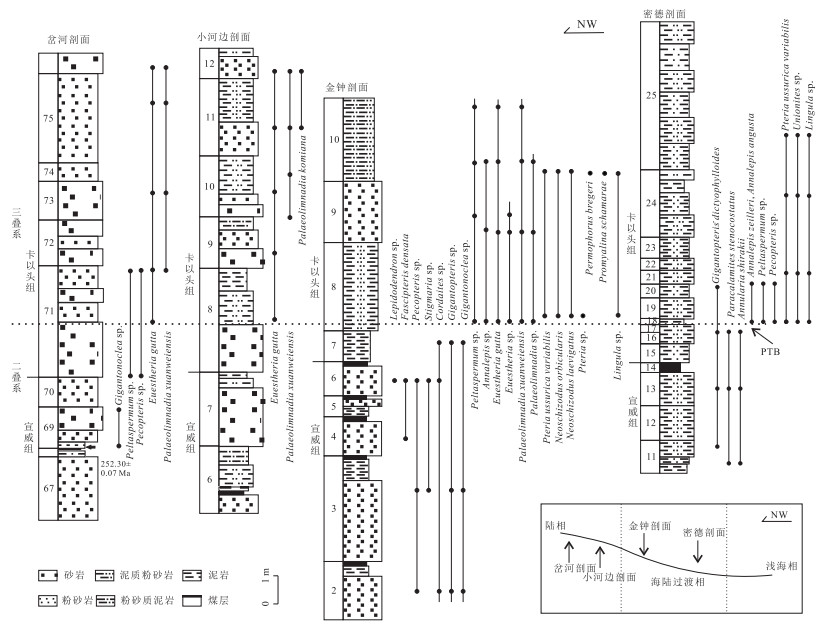
图 3 黔西滇东地区剖面地层对比
岔河剖面(杨逢清等,2005;Shen et al., 2011)、小河边剖面(楚道亮等,2013)、金钟剖面、密德剖面(喻建新等,2008)
Fig. 3. The biostratigraphic correlation of various sections in western Guizhou and eastern Yunnan

图 4 研究区化石标本
a,b.Gigantopteris sp.;a.登记号:JZ06-12.产地:金钟剖面6层顶部;b.登记号:JZ07-40.产地:金钟剖面7层底部;c, d.Pecopteris sp.;c.登记号:JZ06-08.产地:金钟剖面6层顶部;d.登记号:JZ07-48.产地:金钟剖面7层底部;e, f.Annalepis sp.;e.登记号:JZ07-30.产地:金钟剖面8层顶部;f.登记号:JZ07-34.产地:金钟剖面9层顶部;g, h.Peltaspermum sp.;g.登记号:JZ08-28.产地:金钟剖面9层顶部;h.登记号:JZ07-37.产地:金钟剖面8层顶部;i, j.Euestheria gutta Lyutkevich, 1937i.左瓣.登记号:JZ07-01.产地与层位:金钟剖面卡以头组8层顶部;j.右瓣.登记号:JZ08-20.产地:金钟剖面卡以头组9层底部;k, l.Palaeolimnadia xuanweiensis Shen, 1976k.右瓣.登记号:JZ07-04.产地:金钟剖面8层顶部;l.右瓣.登记号:JZ07-05.产地:金钟剖面8层中部;m.Pteria ussurica variabilis Chen and Lan in Gu et al., 1976, 登记号:JZ07-14.产地:金钟剖面8层底部;n.Neoschizodus orbicularis(Bronn, 1837),登记号:JZ07-24.产地:金钟剖面8层底部;o.介壳层, 产地:金钟剖面8层底部;本文研究的化石标本均保存在中国地质大学(武汉)生物地质与环境地质国家重点实验室,采集于贵州威宁金钟剖面宣威组和卡以头组.说明的标本登记号中包括剖面缩写、化石产出层位和标本编号(金钟剖面1层1号标本=JZ01-01).图片使用单反相机(Canon EOS 7D)和体视显微镜(Leica SAP80) 拍摄
Fig. 4. Collection of fossils of research area
表 1 研究区岩性与分层描述
Table 1. Lithology and stratigraphy of research area
东川组(未到顶) 19.紫红色厚层状细砂岩 9.30 m 18.紫红色中厚层状细砂岩 8.20 m 
卡以头组 17.土黄色、紫红色、灰绿色薄层状泥质粉砂岩互层.含叶肢介化石:Euestheria gutta 7.30 m 16.黄绿色-土黄色粉砂质泥岩.含叶肢介化石:Euestheria gutta,Euestheria sp. 11.10 m 15.土黄色厚层状粉砂岩.含植物化石:Annalepis sp.;含叶肢介化石:Euestheria gutta,Euestheria sp.,Palaeolimnadia xuanweiensis,Palaeolimnadia sp. 14.80 m 14.黄绿色薄层粉砂岩夹土黄色粉砂质泥岩.含植物化石:Annalepis sp.,Peltaspermum sp.;含叶肢介化石:Euestheria gutta,Euestheria sp.,Palaeolimnadia xuanweiensis,Palaeolimnadia sp. 9.80 m 13.土黄色薄层粉砂质泥岩.含植物化石:Annalepis sp.,Peltaspermum sp.;含叶肢介化石:Euestheria gutta,Euestheria sp.,Palaeolimnadia xuanweiensis,Palaeolimnadia sp. 5.00 m 12.土黄色薄层粉砂岩.含叶肢介化石:Euestheria gutta,Euestheria sp. 6.20 m 11.土黄色粉砂质泥岩夹黄绿色中薄层砂质泥岩.含植物化石:Annalepis sp.,Peltaspermum sp.;含叶肢介化石:Euestheria gutta,Euestheria sp.,Palaeolimnadia xuanweiensis,Palaeolimnadia sp. 5.30 m 10.黄绿色薄层粉砂质泥岩.含植物化石:Annalepis sp.,Peltaspermum sp.;双壳化石:Pteria ussurica variabilis,Neoschizodus orbicularis,Neoschizodus laevigatus,Promyalina schamarae,Permophorus bregeri,Pteria sp.;叶肢介化石:Euestheria gutta,Euestheria sp.,Palaeolimnadia xuanweiensis,Palaeolimnadia sp.;以及腕足化石Lingula sp.和腹足化石 3.00 m 9.黄绿色薄层粉砂岩.含植物化石:Annalepis sp.;叶肢介化石:Euestheria gutta,Palaeolimnadia xuanweiensis,Palaeolimnadia sp. 2.20 m 8.黄绿色厚层状泥质粉砂岩.含植物化石:Peltaspermum sp.,Annalepis zeilleri,Annalepis angusta;双壳化石Neoschizodus orbicularis,Pteria ussurica variabilis,Neoschizodus laevigatus,Pteria sp.;叶肢介化石:Euestheria gutta,Euestheria sp.,Palaeolimnadia xuanweiensis,Palaeolimnadia sp.;腕足化石Lingula sp. 2.80 m 7.黄绿色薄层状泥岩.含植物化石:Gigantopteris sp.,Cordaites sp.,Gigantonoclea sp. 0.88 m 宣威组(未到底) 6.黄绿色厚层状粉砂岩,下部2层煤线,不连续.含植物化石Pecopteris sp.,Cordaites sp.,Stigmaria sp.,Lepidodendron sp.,Fascipteris densata 1.46 m 5.厚层土黄色泥岩夹厚度不均的煤层 0.60 m 4.黄绿色厚层状粉砂岩,顶部为薄层煤层.含植物化石Gigantopteris sp.,Cordaites sp.,Gigantonoclea sp.,Fascipteris densata 1.60 m 3.灰绿色-土黄色厚层粉砂岩,夹0.70 m白色古土壤,顶部夹0.03 m煤线.含植物化石Gigantopteris sp.,Pecopteris sp.,Gigantonoclea sp.,Stigmaria sp. 3.53 m 2.土黄色厚层状粉砂岩,夹0.38 m灰白色粘土,顶部夹0.08 m煤层.含植物化石Pecopteris sp.,Cordaites sp. 2.41 m 1.土黄色厚层状粉砂岩,可见0.90 m灰白色粘土,带铁质包壳,顶部夹0.06 m煤层.含植物化石Gigantopteris sp.,Cordaites sp.,Gigantonoclea sp.,Pecopteris sp. 未见底  下载: 导出CSV
下载: 导出CSV
-
Alroy, J., Aberhan, M., Bottjer, D.J., et al., 2008.Phanerozoic Trends in the Global Diversity of Marine Invertebrates.Science, 321(5885):97-100.doi: 10.1126/science.1156963 Benton, M.J., 1997.Models for the Diversification of Life.Trends in Ecology & Evolution, 12(12):490-495.doi: 10.1016/S0169-5347(97)84410-2 Bercovici, A., Cui, Y., Forel, M.B., et al., 2015.Terrestrial Paleoenvironment Characterization across the Permian-Triassic Boundary in South China.Journal of Asian Earth Sciences, 98:225-246.doi: 10.1016/j.jseaes.2014.11.016 Burgess, S.D., Samuel, B., Shen, S.Z., 2014.High-Precision Timeline for Earth's Most Severe Extinction.Proceedings of the National Academy of Sciences, 111(9):3316-3321.doi: 10.1073/pnas.1317692111 Cascales-Miñana, B., Diez, J.B., Gerrienne, P., et al., 2015.A Palaeobotanical Perspective on the Great End-Permian Biotic Crisis.Historical Biology, 2015:1-9.doi: 10.1080/08912963.2015.1103237 Chen, J., Tong, J.N., Niu, Z.J., et al., 2010.Lower-Middle Triassic Strata in Qingyan, Guizhou Province, South China.Earth Science, 35(1):51-61(in Chinese with English abstract). http://en.cnki.com.cn/Article_en/CJFDTOTAL-DQKX201001009.htm Chen, J.H., 2005.Macroevolution of Bivalves after the End-Permian Mass Extinction in South China.In:Rong, J.Y., Fang, Z.J., eds., Mass Extinction and Recovery:Evicences from the Palaeozoic and Triassic of South China(Vol.One).University of Science and Technology of China Press, Hefei, 647-700 (in Chinese). Chen, J.H., Yu, J.X., Huang, Q.S., et al., 2011.New Research Progress on the Paleoflora in the Earliest Triassic of Western Guizhouand Eastern Yunnan, South China.Earth Science, 36(3):500-510(in Chinese with English abstract). Chen, Z.Q., Benton, M.J., 2012.The Timing and Pattern of Biotic Recovery Following the End-Permian Mass Extinction.Nature Geoscience, 5(6):375-383.doi: 10.1038/ngeo1475 Chen, Z.Q., George, A.D., Yang, W.R., 2009.Effects of Middle-Late Permian Sea-Level Changes and Mass Extinction on the Formation of the Tieqiao Skeletal Mound in the Laibin Area, South China.Australian Journal of Earth Sciences, 56:745-763.doi: 10.1080/08120090903002581 Chu, D.L., Tong, J.N., Yu, J.X., et al., 2013.The Conchostracan Fauna from the Kayitou Formation of Western Guizhou, China.Acta Palaeontologica Sinica, 52(3):265-268(in Chinese with English abstract). http://en.cnki.com.cn/Article_en/CJFDTOTAL-GSWX201303001.htm Chu, D.L., Tong, J.N., Song, H.J., et al., 2015a.Lilliput Effect in Freshwater Ostracods during the Permian-Triassic Extinction.Palaeogeography, Palaeoclimatology, Palaeoecology, 435:38-52.doi: 10.1016/j.palaeo.2015.06.003 Chu, D.L., Tong, J.N., Song, H.J., et al., 2015b.Early Triassic Wrinkle Structures on Land:Stressed Environments and Oases for Life.Scientific Reports, 5:10109.doi: 10.1038/srep10109 Cui, Y., Bercovici, A., Yu, J., et al., 2015.Carbon Cycle Perturbation Expressed in Terrestrial Permian-Triassic Boundary Sections in South China.Global & Planetary Change, In Press.doi: 10.1016/j.gloplacha.2015.10.018 Erwin, D.H., 1993.The Great Paleozoic Crisis:Life and Death in the Permian.Columbia University Press, New York. Fliche, P., Zeiller, R., 1910.Flore Fossile du Trias en Lorraine et Franche-Comté (Avec Des Considerations Finales Par M.R.Zeiller).Bulletin de la Société des Sciences de Nancy, Ⅲ, Ⅺ 1:222-286. Gastaldo, R.A., Kamo, S.L., Neveling, J., et al., 2015.Is the Vertebrate-Defined Permian-Triassic Boundary in the Karoo Basin, South Africa, the Terrestrial Expression of the End-Permian Marine Event?Geology, 43(10):939-942.doi: 10.1130/G37040.1 Grauvogel-Stamm L., Lugardon, B., 2001.The Triassic Lycopsids Pleuromeia and Annalepis:Relationship, Evolution and Origin.American Fern Journal, 91:115-149.doi: 10.1640/0002-8444(2001)091[0115:TTLPAA]2.0.CO;2 Hou, J.P., Ouyang, S., 2000.Palynoflora from the Sunjiagou Formation in Liulin County, Shanxi Province.Acta Palaeontologica Sinica, 39(3):356-368(in Chinese with English abstract). http://en.cnki.com.cn/Article_en/CJFDTOTAL-GSWX200003005.htm Huang, Y.F., Tong, J.N., 2014.Advance in the Study of the Permian-Triassic Bivalves.Advances in Earth Science, 29(8):922-933(in Chinese with English abstract). Huang, Y.F., Tong, J.N., Xiang, Y., et al., 2015.The Extinction and Delayed Recovery of Bivalves during the Permian-Triassic Crisis.Earth Science, 40(2):334-345(in Chinese with English abstract). Jiang, H.S., Lai, X.L., Luo, G.M., et al., 2007.Restudy of Conodont Zonation and Evolution across the P/T Boundary at Meishan Section, Changxing, Zhejiang, China.Global & Planetary Change, 55(1-3):39-55.doi: 10.1016/j.gloplacha.2006.06.007 Komatsu, T., Huyen, D.T., Chen, J.H., 2008.Lower Triassic Bivalve Assemblages after the End-Permian Mass Extinction in South China and North Vietnam.Paleontological Research, 12(2):119-128.doi: 10.2517/1342-8144(2008)12[119:LTBAAT]2.0.CO;2 Kozur, H.W., Weems, R.E., 2010.The Biostratigraphic Importance of Conchostracans in the Continental Triassic of the Northern Hemisphere.Geological Society, London, Special Publications, 34(1):315-417.doi: 10.1144/SP334.13 Kozur, H.W., Weems, R.E., 2011.Detailed Correlation and Age of Continental Late Changhsingian and Earliest Triassic Beds:Implications for the Role of the Siberian Trap in the Permian-Triassic Biotic Crisis.Palaeogeography Palaeoclimatology Palaeoecology, 308(1-2):22-40.doi: 10.1016/j.palaeo.2011.02.020 Li, X.X., 1995.Fossil Floras of China through the Geological Ages.Guangdong Science and Technology Press, Guangdong, 229 -253(in Chinese). Liu, S.W., Li, Y.G., Wang, S.T., et al., 1990.Study of Some Permian-Triassic Fauna Fossils in Duwa, Hotan, Xinjiang.Xinjiang Geology, 8(1):36-46(in Chinese with English abstract). Liu, X., Gituru, W.R., Wang, Q.F., 2004.Distribution of Basic Diploid and Polyploid Species of Isoetes in East Asia.Journal of Biogeography, 31:1239-1250.doi: 10.1111/j.1365-2699.2004.01115.x Looy, C.V., Brugman, W.A., Dilcher, D.L., et al., 1999.The Delayed Resurgence of Equatorial Forests after the Permian-Triassic Ecologic Crisis.Proceedings of the National Academy of Sciences, 96(24):13857-13862.doi: 10.1073/pnas.96.24.13857 Looy, C.V., Twitchett, R.J., Dilcher, D.L., et al., 2001.Life in the End-Permian Dead Zone.Proceedings of the National Academy of Sciences, 98(14):7879-7883.doi: 10.1073/pnas.131218098 Lucas, S.G., 1998.Global Triassic Tetrapod Biostratigraphy and Biochronology.Palaeogeography Palaeoclimatology Palaeoecology, 143(98):347-384.doi: 10.1016/S0031-0182(98)00117-5 Lucas, S.G., 2009.Timing and Magnitude of Tetrapod Extinctions across the Permo-Triassic Boundary.Journal of Asian Earth Sciences, 36(6):491-502.doi: 10.1016/j.jseaes.2008.11.016 Mcroberts, C.A., 2010.Biochronology of Triassic Bivalves.Geological Society London Special Publications, (1):201-219.doi: 10.1144/SP334.9 Meng, F.S., 1994.Discovery of Pleuromeia-Annalepis Flora in South China and Its Significance.Chinese Science Bulletin, 39:130-134. Meng, F.S., Zhang, Z.L., Niu, Z.J., et al., 2000.Primitive Lycopsid Flora in the Yangtze Valley of China and Systematics and Evolution of Isoetales.Hunan Science and Technology Press, Changsha (in Chinese with English abstract). Nakazawa, K., 1981.Permian and Triassic Bivalves of Kashmir.In:Nakazawa, K., Kapoor, H.M., eds., The Upper Permian and Lower Triassic Faunas of Kashmir, Palaeontologica Indica, New Series, 46, 87-122. Ouyang, S., 1991.Transitional Palynofloras from Basal Lower Triassic of China and Their Ecological Implications, with Special Reference to Paleophyte/Mesophyte Problems.In:Jin Y.G., ed., Palaeoecology of China 1, Nanjing University Press, Nanjing, 168-196. Pang, Q.Q., Jin, X.C., 2004.Ostracoda of the Guodikeng Formation and the Continental Permo Triassic Boundary of the Dalongkou Section, Jimsar, Xinjiang.Professional Papers of Stratigraphy and Palaeontology, 00:205-246(in Chinese with English abstract). Peng, Y.Q., Tong, J.N., 1999.The Study on Permian-Triassic Boundary Bed in Yangtze Platform.Earth Science, 24(1):39-48(in Chinese with English abstract). Peng, Y.Q., Yin, H.F., Yang, F.Q., 2001.Advance in the Study of Terrestrial Permian-Triassic Boundary.Advance in Earth Science, 16(6):769-776(in Chinese with English abstract). http://en.cnki.com.cn/Article_en/CJFDTotal-DXJZ200106007.htm Peng, Y.Q., Zhang, S.X., Yu, T.X., et al., 2005.High-Resolution Terrestrial Permian-Triassic Eventostratigraphic Boundary in Western Guizhou and Eastern Yunnan, Southwestern China.Palaeogeography Palaeoclimatology Palaeoecology, 215(3):285-295.doi: 10.1016/j.palaeo.2004.09.009 Posenato, R., 2009.Survival Patterns of Macrobenthic Marine Assemblages during the End-Permian Mass Extinction in the Western Tethys (Dolomites, Italy).Palaeogeography, Palaeoclimatology, Palaeoecology, 280(1):150-167.doi: 10.1016/j.palaeo.2009.06.009 Raup, D.M., 1979.Size of the Permo-Triassic Bottleneck and Its Evolutionary Implications.Science, 206(4415):217-218.doi: 10.1126/science.206.4415.217 Retallack, G.J., 1975.The Life and Times of a Triassic Lycopod.Alcheringa, 1:3-29.doi: 10.1080/03115517508619477 Retallack, G.J., 1995.Permian-Triassic Life Crisis on Land.Science, 267(5194):77-80.doi: 10.1126/science.267.5194.77 Retallack, G.J., 1997.Earliest Triassic Origin of Isoetes and Quillwort Evolutionary Radiation.Journal of Paleontology, 71:500-521.doi: 10.1017/S0022336000039524 Schneebeli-Hermann, E., Bucher, H., 2015. Palynostratigraphy at the Permian-Triassic Boundary of the Amb Section, Salt Range, Pakistan.Palynology, 39(1):1-18.doi: 10.1080/01916122.2014.921648 Scholze, F., Golubev, V.K., Niedwiedzki, G., et al., 2015.Early Triassic Conchostracans (Crustacea:Branchiopoda) from the Terrestrial Permian-Triassic Boundary Sections in the Moscow Syncline.Palaeogeography Palaeoclimatology Palaeoecology, 429:22-40.doi: 10.1016/j.palaeo.2015.04.002 Sephton, M.A., Jiao, D., Engel, M.H., et al., 2015.Terrestrial Acidification during the End-Permian Biosphere Crisis? Geology, 43(2):159-162.doi: 10.1130/G36227.1 Shen, S.Z., He, X.L., Shi, G.R., 1995.Biostratigraphy and Correlation of Several Permian-Triassic Boundary Sections in Southwestern China.Journal of Southeast Asian Earth Sciences, 12(1):19-30.doi: 10.1016/0743-9547(95)00026-7 Shen, S.Z., Henderson, C.M., Bowring, S.A., et al., 2010.High-Resolution Lopingian (Late Permian) Timescale of South China.Geological Journal, 45(2-3):122-134.doi: 10.1002/gj.1232 Shen, S.Z., Crowley, J.L., Wang, Y., et al., 2011.Calibrating the End-Permian Mass Extinction.Science, 334(6061):1367.doi: 10.1126/science.1213454 Shen, W.J., Sun, Y.G., Lin, Y.T., et al., 2011.Evidence for Wildfire in the Meishan Section and Implications for Permian-Triassic Events.Geochimica et Cosmochimica Acta, 75(7):1992-2006.doi: 10.1016/j.gca.2011.01.027 Song, H.J., Wignall, P.B., Tong, J.N., et al., 2013.Two Pulses of Extinction during the Permian-Triassic Crisis.Nature Geoscience, 6(1):52-56.doi: 10.1038/ngeo1649 Tian, L., Tong, J.N., Song, H.J., et al., 2014.Foraminiferal Evolution and Formation of Oolitic Limestone near Permian-Triassic Boundary at Yangou Section, Jiangxi Province.Earth Science, 39(11):1473-1486(in Chinese with English abstract). http://en.cnki.com.cn/Article_en/CJFDTotal-DQKX201411001.htm Tian, Y.T., Yu, J.X., Feng, Q.L., 2008.The Discovery of Ophiceras in the Kayitou Formation in Eastern Yunnan Province and Its Signification.Journal of Stratigraphy, 32(2):153-158(in Chinese with English abstract). Tong, J.N., Yin, H.F., 2015.Triassic Chronostratigraphy and Chinese Stages.Earth Science, 40(2):189-197(in Chinese with English abstract). http://en.cnki.com.cn/Article_en/CJFDTOTAL-DQKX201502001.htm Tu, C.Y., Chen, Z.Q., Retallack, G.J., et al.2016.Proliferation of MISS-Related Microbial Mats Following the End-Permian Mass Extinction in Terrestrial Ecosystems:Evidence from the Lower Triassic of the Yiyang Area, Henan Province, North China.Sedimentary Geology, 333:50-69.doi: 10.1016/j.sedgeo.2015.12.006 Wang, S.Y., Yin, H.F., 2001a.Study on Terrestrial Permian-Triassic Boundary in Eastern Yunnan and Western Guizhou.China University of Geosciences Press, Wuhan (in Chinese). Wang, S.Y., Yin, H.F., 2011b.Advance in the Study of the Terrestrial Permian-Triassic Boundary, South China.Chineses Geology, 28(7):16-21(in Chinese). Wang, Z.Q., 1996.Recovery of Vegetation from the Terminal Permian Mass Extinction in North China.Review of Palaeobotany & Palynology, 91(1):121-142.doi: 10.1016/0034-6667(95)00069-0 Wang, Z.Q., Chen, A.S., 2001.Traces of Arborescent Lycopsids and Dieback of the Forest Vegetation in Relation to the Terminal Permian Mass Extinction in North China.Review of Palaeobotany & Palynology, 117(4):217-243.doi: 10.1016/S0034-6667(01)00094-X Ward, P.D., Montgomery, D.R., Smith, R., 2000.Altered River Morphology in South Africa Related to the Permian-Triassic Extinction.Science, 289(5485):1740-1743.doi: 10.1126/science.289.5485.1740 Yang, F.Q., Yin, H.F., Yu, J.X., et al., 2005.Study of the Terrestrial Permian-Triassic Boundary of the Chahe Section, Weining, Guizhou Province.Science in China(Ser.D), 35(6):519-529 (in Chinese). Yang, H., Zhang, S.X., Jiang, H.S., et al., 2006.Age and General Characteristics of Calcimicrobialite near the Permian-Triassic Boundary in Chongyang, Hubei Province.Earth Science, 31(2):165-170(in Chinese with English abstract). Yang, Z.Y., Yin, H.F., Wu, S.B., et al., 1987.Permian-Triassic Boundary Stratigraphy and Fauna of South China.Geological Publishing House, Beijing (in Chinese with English abstract). Yao, Z.Q., 1978.On the Age of " Gigantopteris Coal Series" and Gigantopteris-Flora in South China.Acta Palaeontologica Sinica, 17(1):81-89 (in Chinese with English abstract). Yao, Z.Q., Xu, J.T., Zhen, Z.G., et al., 1980.Biostratigraphy of Late Permian in West Guizhouand and Easte Yunnan and the Permo-Triassic Boundary.In:Nanjing Institute of Geology and Palaeontology, Academia Sinica, ed., Stratigraphy and Palaeontology of Late Permian Coal-Bearing Formations.Science Press, Beijing (in Chinese). Ye, M.N., 1979.On Some Middle Triassic Plants from Hupeh and Szechuan.Acta Palaeontologica Sinica, 18(1):73-82(in Chinese with English abstract). Yin, H.F., Xie, S.C., Luo, G.M., et al., 2012.Two Episodes of Environmental Change at the Permian-Triassic Boundary of the GSSP Section Meishan.Earth-Science Reviews, 115(3):163-172.doi: 10.1016/j.earscirev.2012.08.006 Yin, H.F., Zhang, K.X., Tong, J.N., et al., 2001.The Global Stratotype Section and Point (GSSP) of the Permian-Triassic Boundary.Episodes, 24(2):102-114. http://ci.nii.ac.jp/naid/10020538786 Yu, J.X., Broutin, J., Chen, Z.Q., et al., 2015.Vegetation Changeover across the Permian-Triassic Boundary in Southwest China:Extinction, Survival, Recovery and Palaeoclimate:A Critical Review.Earth-Science Reviews, 149:203-224.doi: 10.1016/j.earscirev.2015.04.005 Yu, J.X., Huang, Q.S., Broutin, J., 2008.The Early Early Triassic Annalepis from Western Guizhou and Eastern Yunnan, South China.Acta Palaeontologica Sinica, 47(3):292-300(in Chinese with English abstract). http://en.cnki.com.cn/Article_en/CJFDTOTAL-GSWX200803006.htm Zhang, H., Cao, C.Q., Liu, X.L., et al., 2015.The Terrestrial End-Permian Mass Extinction in South China.Palaeogeography, Palaeoclimatology, Palaeoecology, In Press.doi: 10.1016/j.palaeo.2015.07.002 Zhang, W.T., Chen, P.J., Shen, Y.B., 1976.The Conchostracan of China.Science Press, Beijing(in Chinese). 陈建华, 喻建新, 黄其胜, 等, 2011.黔西滇东地区早三叠世早期植物化石研究的新进展.地球科学, 36(3):500-510. http://earth-science.net/WebPage/Article.aspx?id=2113 陈晶, 童金南, 牛志军, 等, 2010.贵州青岩地区早-中三叠世生物地层.地球科学, 35(1):51-61. http://earth-science.net/WebPage/Article.aspx?id=1929 陈金华, 2005.华南二叠纪末大灭绝后双壳类的宏演化阶段.见戎嘉余, 方宗杰, 生物大绝灭与复苏:来自华南古生物代和三叠纪的证据(上卷).合肥:中国科学技术大学出版社, 647-700. 楚道亮, 童金南, 喻建新, 等.2013.黔西地区卡以头组叶肢介动物群.古生物学报, 52(3):265-280. http://www.cnki.com.cn/Article/CJFDTOTAL-GSWX201303001.htm 侯静鹏, 欧阳舒, 2000.山西柳林孙家沟组孢粉植物群.古生物学报, 39(3):356-368. http://www.cnki.com.cn/Article/CJFDTOTAL-GSWX200003005.htm 黄云飞, 童金南, 2014.古-中生代之交双壳类演变研究进展.地球科学进展, 29(08):922-933. http://www.cnki.com.cn/Article/CJFDTOTAL-DXJZ201408008.htm 黄云飞, 童金南, 向烨, 等, 2015.二叠纪-三叠纪之交双壳类的灭绝与复苏过程.地球科学, 40(2):334-345. http://earth-science.net/WebPage/Article.aspx?id=3048 李星学, 1995.中国地质时期植物群.广东:广东科技出版社, 229-253. 刘淑文, 李友桂, 王士涛, 等.1990.新疆和田杜瓦二叠-三叠纪动物化石研究.新疆地质, 8(1):36-46. http://www.cnki.com.cn/Article/CJFDTOTAL-XJDI199001004.htm 孟繁松, 张振来, 牛志军, 等, 2000.长江流域原始石松植物及水韭目植物分类与演化.长沙:湖南科学技术出版社. 庞其清, 金小赤, 2004.新疆吉木萨尔大龙口锅底坑组的介形虫及陆相二叠系-三叠系界线.地层古生物论文集, 00:205-246. http://cpfd.cnki.com.cn/Article/CPFDTOTAL-ZGDJ200400008015.htm 彭元桥, 童金南, 1999.扬子台区二叠-三叠系界线层综合地层学研究.地球科学, 24(1):39-48. http://earth-science.net/WebPage/Article.aspx?id=744 彭元桥, 殷鸿福, 杨逢清, 2001.陆相二叠系-三叠系界线研究进展.地球科学进展, 16(6):769-776. http://www.cnki.com.cn/Article/CJFDTOTAL-DXJZ200106007.htm 田力, 童金南, 宋海军, 等, 2014.江西乐平沿沟二叠纪末有孔虫的演变及对鲕状灰岩的古环境指示意义.地球科学, 39(11):1473-1486. http://earth-science.net/WebPage/Article.aspx?id=2975 田云涛, 喻建新, 冯庆来, 2008.Ophiceras(蛇菊石)在滇东卡以头组的发现及其意义.地层学杂志, 32(2):153-158. http://www.cnki.com.cn/Article/CJFDTOTAL-DCXZ200802003.htm 童金南, 殷鸿福, 2015.三叠纪年代地层与中国建阶.地球科学:中国地质大学学报, 40(2):189-197. http://www.cnki.com.cn/Article/CJFDTOTAL-DQKX201502001.htm 王尚彦, 殷鸿福, 2001a.滇东黔西陆相二叠纪-三叠纪界线地层研究.武汉:中国地质大学出版社. 王尚彦, 殷鸿福, 2001b.华南陆相二叠-三叠系界线地层研究新进展.中国地质, 28(7):16-21. http://www.cnki.com.cn/Article/CJFDTOTAL-DIZI200107002.htm 杨逢清, 殷鸿福, 喻建新, 等, 2005.贵州威宁岔河陆相二叠系-三叠系界线地层研究.中国科学(D辑), 35(6):519-529. http://www.cnki.com.cn/Article/CJFDTOTAL-JDXK200506004.htm 杨浩, 张素新, 江海水, 等, 2006.湖北崇阳二叠纪-三叠纪之交钙质微生物岩的时代及基本特征.地球科学, 31(2):165-170. http://earth-science.net/WebPage/Article.aspx?id=1573 杨遵仪, 殷鸿福, 吴顺宝, 等, 1987.华南二叠-三叠系界线地层及动物群.北京:地质出版社. 姚兆奇, 1978.华南"大羽羊齿煤系"和大羽羊齿植物群的时代.古生物学报, 17(1):81-89. http://www.cnki.com.cn/Article/CJFDTOTAL-GSWX197801004.htm 姚兆奇, 徐均涛, 郑灼官, 等, 1980.黔西滇东晚二叠世生物地层和二叠系与三叠系的界线问题.见黔西滇东晚二叠世含煤地层和古生物群.北京:科学出版社. 叶美娜, 1979.湖北、四川中三叠统植物化石的发现.古生物学报, 18(1):73-82. http://www.cnki.com.cn/Article/CJFDTOTAL-GSWX197901004.htm 喻建新, 黄其胜, Broutin, J., 等, 2008.黔西滇东早三叠世早期Annalepis(脊囊属)的出现及其地层意义.古生物学报, 47(3):292-300. http://www.cnki.com.cn/Article/CJFDTOTAL-GSWX200803006.htm 张文堂, 陈丕基, 沈炎彬, 1976.中国的叶肢介化石.北京:科学出版社. -
 点击查看大图
点击查看大图
计量
- 文章访问数: 4574
- HTML全文浏览量: 2087
- PDF下载量: 51
- 被引次数: 0
